



Introduction
Rare occurrences of Middle Pleistocene marine faunas are known from some outcrops and boreholes in the southern North Sea Basin, yet their stratigraphic context is poorly understood (Meijer & Cleveringa, Reference Meijer and Cleveringa2009). Therefore, the marine conditions (temperature regimes, biodiversity and palaeogeography) in the North Sea Basin are very poorly understood for that time interval. In the Netherlands, north of the maximum extension limit of the Drenthe Glacial Substage ice sheet (Late Saalian, marine isotope stage MIS 6), such deposits have often been eroded and sometimes deformed, whereas south of this limit only very few, isolated sites exist. Although some borehole records of mollusc-bearing marine Middle Pleistocene intervals have been reported (Ter Wee, Reference Ter Wee1976; Meijer & Cleveringa, Reference Meijer and Cleveringa2009), fossil recovery is poor and detailed age control is mostly lacking. This has limited our understanding of the evolution of the North Sea Basin during the Middle Pleistocene and makes comparison with Late Pleistocene environmental conditions impossible. For example, the timing of marine connections through the English Channel and the nature of marine settings in successive interglacial intervals are still unclear (Gibbard et al., Reference Gibbard, West, Zagwijn, Balson, Burger, Funnell, Jeffery, de Jong, van Kolfschoten, Lister, Meijer, Norton, Preece, Rose, Stuart, Whiteman and Zalasiewicz1991; Hijma et al., Reference Hijma, Cohen, Roebroeks, Westerhoff and Busschers2012). In order to address these issues, records are needed from marine Middle Pleistocene intervals with faunas that are chronologically well-constrained.
Borehole B01H0189 (Hollum, Ameland, the Netherlands; Fig. 1) provides a unique succession of three marine mollusc associations that has the potential to be a key section to document and understand marine interglacial conditions in the North Sea Basin during the late Middle Pleistocene to Holocene high stands. Amino acid racemisation (AAR) data from the borehole show that the lower marine interval is of late Middle Pleistocene MIS7 age (Meijer & Cleveringa, Reference Meijer and Cleveringa2009), thereby constraining the upper two marine intervals to Late Pleistocene (MIS5e) and Holocene (MIS1) high stands. The aim of this paper is to document and characterise the fauna composition and diversity in order to compare climate and environmental conditions in shallow marine settings during successive marine (interglacial) high stands in the southern North Sea Basin since the late Middle Pleistocene.
Fig. 1. Location of borehole B01H0189 Hollum.
Materials and methods
Borehole B01H0189 is a continuously cored borehole located at RD X173000/Y605400 (53.4346° N, 5.6580° E) with a surface level at +1.51 m NAP (Dutch Ordnance Datum). The core was drilled in October 1994 and reached a depth of 40.0 m below surface (b.s.). The core was obtained by a mechanical bailer-drilling unit that uses a steel-cased borehole and a steel sediment core sampler, yielding undisturbed samples in PVC tubes of 10 cm diameter and 1 m length (Busschers et al., Reference Busschers, Kasse, Van Balen, Vandenberghe, Cohen, Weerts, Wallinga, Johns, Cleveringa and Bunnik2007). The cores were split and one half was used to produce standardised lithological descriptions and photographs at TNO-NITG (now TNO – Geological Survey of the Netherlands) by M.A. Smakman. Down-hole contamination, only sporadically occurring at the 1 m core segment transitions, was excluded from further sampling and analysis. The borehole data are available at https://www.dinoloket.nl/en (referred to as dinoloket below). The detailed core photographs were used to cross-check and refine descriptions and especially identify sedimentary structures following Van den Berg & Nio (Reference Van den Berg and Nio2010). The reported AAR age estimates derive from Meijer & Cleveringa (Reference Meijer and Cleveringa2009) who used methods outlined in Bowen et al. (Reference Bowen, Hughes, Sykes and Miller1989) and Bowen (Reference Bowen, Goodfriend, Collins, Fogel, Macko and Wehmiller2000). The other half of the core was sampled for analysis of pollen content (Veldkamp & Cleveringa, Reference Veldkamp and Cleveringa1997), diatoms (De Wolf, Reference De Wolf1996), foraminifera (Van Leeuwen, Reference Van Leeuwen1996) and molluscs (Pouwer & Meijer, Reference Pouwer and Meijer1996). Palynological and micropalaeontological samples were taken in series from fine-grained (clayey) and peaty intervals with sampling resolutions of 10 to 50 cm. These results were presented in internal reports and have not been published before. We reanalysed the molluscs in order to study the succession. In total, 39 samples were analysed: typically a sample weighted 150 g. After wet-sieving (minimum mesh width 0.5 mm) and drying, samples were picked and species identified. Unique parts of shells (hinges of bivalves and apertures of gastropods) were counted, as were recognisable fragments of uncommon species. A number of qualitative criteria were used to determine whether a shell or fragment could represent an in situ record. Most important are lack of abrasion (on edges or on umbones), preservation of fine surface details and lack of discoloration and bioerosion.
There are some taxonomic uncertainties in our material. Bittium reticulatum concerns a species complex that may also involve B. scabrum, B. latreilli and/or B. jadertinum (Wesselingh et al., Reference Wesselingh, Rijken, van Nieulande, Janse and Pouwer2012). As these species can only be identified reliably with adult specimens and B. reticulatum in general is the dominating species in the Late Quaternary records of the Netherlands, we have assigned all Bittium to that species (Wesselingh et al., Reference Wesselingh, Rijken, van Nieulande, Janse and Pouwer2012). We are uncertain about the taxonomic status of Pussilina radiata balkeii, which is the subject of further study. Tritia reticulata s.l. may include the similar T. nitida because these species can only be reliably identified on adult material, hence their grouping (Rijken & Pouwer, Reference Rijken and Pouwer2014). The same applies to Stagnicola palustris s.l., for which several species are known that are difficult or impossible to distinguish on limited shell characteristics. The unidentified Nucula specimens must be either Nucula nitidosa or N. nucleus. The former is well-known from Holocene deposits, and the latter was very common during MIS5e in the central Netherlands. Identification is only possible when fully preserved valves of adult specimens are available (Moerdijk et al., Reference Moerdijk, Janssen, Wesselingh, Peeters, Pouwer, van Nieulande, Janse, van der Slik, Meijer, Rijken, Cadée, Hoeksema, Doeksen, Bastemeijer, Strack, Vervoenen and ter Poorten2010). Mactra stultorum s.l. may include the subspecies Mactra stultorum plistoneerlandica for pre-Holocene intervals (see Moerdijk et al., Reference Moerdijk, Janssen, Wesselingh, Peeters, Pouwer, van Nieulande, Janse, van der Slik, Meijer, Rijken, Cadée, Hoeksema, Doeksen, Bastemeijer, Strack, Vervoenen and ter Poorten2010, for details). Here we use the name Macoma balthica instead of Limecola balthica as we consider the proposed genus assignment by Huber et al. (Reference Huber, Langleit and Kreipl2015) not to be supported by sufficient evidence. Due to the taxonomic uncertainties, our estimates of specimen numbers are somewhat conservative. Salinity preferences as well as the broad temperature/biogeographic character of the mollusc species were assembled from De Bruyne et al. (Reference De Bruyne, van Leeuwen, Gmelig Meyling and Daan2013), Cleveringa et al. (Reference Cleveringa, Meijer, van Leeuwen, de Wolf, Pouwer, Lissenberg and Burger2000) and Peacock (Reference Peacock1993). Multivariate analyses (non-metric multidimensional scaling (NMDS), detrended correspondence analyses (DCA) and cluster analyses) were performed using PAST (Hammer et al., Reference Hammer, Harper and Ryan2001) version 4.01 (https://folk.uio.no/ohammer/past/) to investigate similarities of samples within assemblages and characterise the different assemblages.
Although the paper focuses on molluscs, an exception is made for the remains of the tubes of the polychaete genus Pectinaria. These tubes are built of sand grains glued together with mucus. This fabric does not survive ages older than the Holocene, therefore the presence of Pectinaria is an excellent indicator for a Holocene age in a marine environment.
Lithological units and palaeoenvironmental interpretations
Five lithological units are distinguished that can be attributed to the Naaldwijk, Eem, Boxtel and Urk Formations (Fig. 2). Although our lithostratigraphic terminology follows the nomenclature of TNO-GSN (2020), the assignment of geological units below the Eem Formation to the Boxtel and Urk Formation in the study core, as well as from cores depicted in Figure 5, differs from Dinoloket (see Discussion).

Fig. 2. Lithostratography and palaeoenvironmental interpretation of borehole B01H0189 Hollum (depth in core sections are in m b.s.). AAR-C indicates the level (35.12–35.22 m b.s.) from where Macoma balthica and Spisula subtruncata provided AAR ratios that were attributed to AAR zone C of Meijer & Cleveringa (Reference Meijer and Cleveringa2009). Core photographs show intervals with representative lithologies.
Unit 1 (39.00–40.00 m b.s.) (Urk Formation, Tijnje or Veenhuizen Member)
Unit 1 consists of decalcified silty clay. The colour of the clay changes upwards from brownish to dark grey. The entire interval is penetrated by vertical mm-thick structures that probably represent roots. At the top of the unit a c.2 cm thick organic layer is present consisting of peat or a wood fragment. This layer likely reflects a preserved basal part of a thicker peat layer that was eroded afterwards. Neither molluscs nor diatoms were recovered.
Unit 1 is dominated by Quercus and Corylus (in lower amounts compared to the overlying Unit 2) and, compared to that unit, relatively high amounts of Alnus. Carpinus and Taxus are lacking and indications for reworking are common. The palynological content of Unit 1 indicates a late Middle Pleistocene interglacial (Zagwijn, Reference Zagwijn1973, Reference Zagwijn and Turner1996a; De Jong, Reference De Jong1988). We interpret Unit 1 to have formed in a flood basin environment with soil formation. The thin organic layer at the top of the unit shows the flood basin likely changed into a swamp environment.
Unit 2 (32.80–39.00 m b.s.) (Urk Formation, Tijnje Member)
Unit 2 consists of grey very fine-grained shelly sands and clays. The clay layers strongly dominate the unit and generally show evidence for bioturbation, especially in the upper part. Between c.39.00 and 35.50 m b.s. the clay layers range in thickness from 1 to 10 cm while higher up the unit the clay becomes much more massive, with only one c.0.2 m thick sand intercalation occurring around −33.90 m b.s. The mollusc fauna is dominated by Abra alba and Spisula subtruncata. No diatoms were recovered.
Unit 2 is dominated by Quercus, Corylus, Carpinus, Pinus, Picea and Alnus pollen with low amounts of Taxus and heathland pollen, although reworking is significant. We regard the pollen as indicative for a late Middle Pleistocene interglacial (Zagwijn, Reference Zagwijn1973, Reference Zagwijn and Turner1996a; De Jong, Reference De Jong1988).
Two AAR ratios have previously been obtained from this assemblage. A specimen of Macoma balthica from sample 35.12–35.22 m b.s. yielded a AAR ratio of 0.32, whereas a specimen of Spisula subtruncata from the same sample yielded a AAR ratio of 0.27 (Meijer & Cleveringa, Reference Meijer and Cleveringa2009). This puts them firmly in the Amino Zone C of these authors. By correlation to dated intervals within the same and overlying and underlying amino zones in boreholes throughout the southern North Sea Basin they inferred an MIS7 age for their Amino Zone C that corresponds to the Oostermeer Interglacial of Meijer (Reference Meijer2003).
The sharp sand–clay transitions and presence of clay in the system, together with the marine mollusc species, strongly suggest Unit 2 was deposited in a tidal environment, probably in a tidal flat.
Unit 3 (c.30.40–32.80 m b.s. (Boxtel Formation)
Unit 3 is generally characterised by a high organic content. The lower part of the unit between 32.80 and 32.05 m b.s. consists of medium- and coarse-grained sands, sometimes alternating with thin clay- or detritus layers. The lower 30 cm of this interval is characterised by coarse sand with sandstone and flint gravels. This coarse layer, which can be traced laterally throughout other cores in the area where it also contains Scandinavian crystalline material (e.g. B01H0098; Fig. 3), is interpreted as an erosive residue of former glacial sediments. Between 32.05 and 30.55 m b.s. the unit is dominated by dark-brownish to dark-grey very fine sands with mica, alternating with detritus layers. Thin clay layers occur in the lower part of this interval. Between 30.55 and 30.50 m b.s. the unit consists of humic clay. The upper c.0.5 m of Unit 3 is decalcified. The local (non-Rhine) nature of these sediments makes this interval formally part of the Boxtel Formation, although future study should explore this stratigraphic assignment in more detail. The interval 30.50–30.40 m b.s. was not recovered in the core.
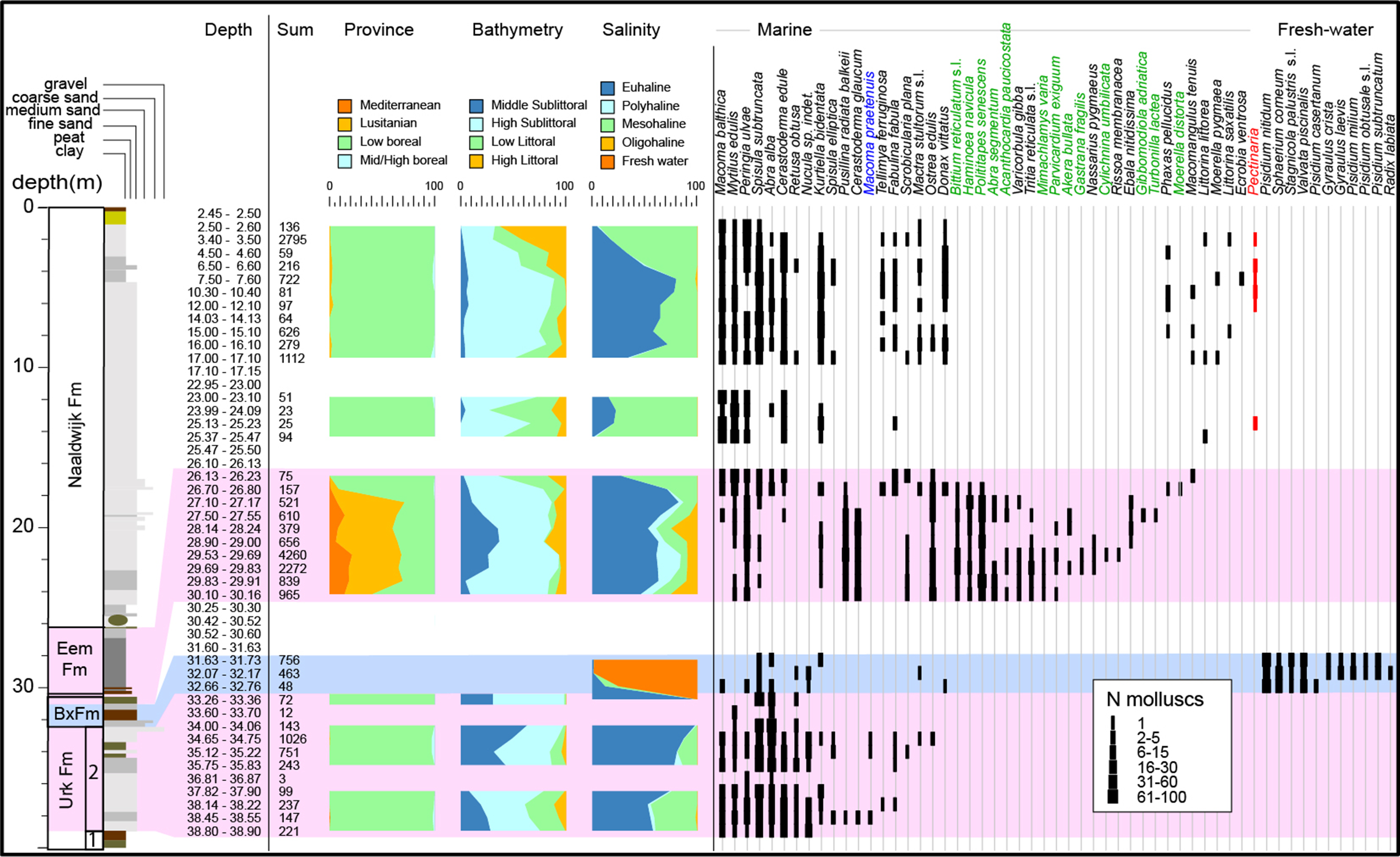
Fig. 3. Mollusc data from borehole B01H0189 Hollum. Only selected species shown. Sum = number of counted specimens. Species in green represent typical Eemian species; in blue is a pre-Eemian species. In red the tubeworm Pectinaria is shown that is a typical indicator of Holocene marine settings. Raw data in the Supplementary Material available online at https://doi.org/10.1017/njg.2021.2.
The sands in this unit contain variable amounts of molluscs dominated by thermophile fresh water species; diatoms are absent. The minor marine component resembles the fauna from the unit below and we interpret the molluscs to be the result of reworking (e.g. by bioturbation).
Unit 3 is dominated by Quercus, Corylus, Pinus, Picea, Betula and Alnus pollen and low amounts of heathland pollen. Remarkable is the occurrence of Juniperus, Equisetum, Cyperaceae and Selaginella pollen and spores. We regard the palynological assemblage as reflecting a freshwater pioneer vegetation. Since the amount of reworked pre-Quaternary and Quaternary pollen is relatively high, and the amount of tree pollen varies strongly, we argue that at least part of the assemblage resulted from reworking. The pollen data are not diagnostic for a specific age. However, the position above (reworked) glacial sediments of the Drenthe Formation and below the Eemian sediments of Unit 4 (see below) makes a latest Late Saalian (MIS6) age most likely. Future study should focus on possible links to other Late Saalian pollen records that were described in, for example, the IJssel Valley basin (Busschers et al., Reference Busschers, Kasse, Van Balen, Vandenberghe, Cohen, Weerts, Wallinga, Johns, Cleveringa and Bunnik2007).
The alternation of fine-grained, mica-rich sands and organic clays with debris and the presence of a freshwater pioneer vegetation suggests Unit 3 was deposited in a low-energy fluvial or lake environment.
Unit 4 (26.24–30.40 m b.s.) (Eem Formation)
Between 26.24 and 30.16 m b.s., Unit 4 is dominated by grey shell-bearing clays that alternate with silt layers. Sand content increases upward, with clear sand layers present above c.27 m b.s. Between 29.50 and 29.95 m b.s. a thick mollusc concentrate is present. A species-rich, very well-preserved marine mollusc fauna containing a number of thermophile Eemian indicator species (Spaink, Reference Spaink1958; Cleveringa et al., Reference Cleveringa, Meijer, van Leeuwen, de Wolf, Pouwer, Lissenberg and Burger2000; Van Leeuwen et al., Reference Van Leeuwen, Beets, Bosch, Burger, Cleveringa, van Harten, Herngreen, Kruk, Langereis, Meijer, Pouwer and de Wolf2000) (including Bittium reticulatum s.l., Acanthocardia paucicostata, Polititapes senescens, Haminoea navicula) indicates the unit is of Eemian age. The dominance of Bittium reticulatum s.l. and small rissoid gastropods implies possibly seagrass/seaweed environments, and the abundance of Cerastoderma glaucum shows somewhat depressed (mesohaline–polyhaline) salinity settings. The diatom association represents a marine–littoral Eemian association, and the occurrence of epiphytic and epipsammic species is indicative of seaweed meadows. The pollen spectra between 27.50 and 30.16 m b.s. represent Eemian zone E4 (due to the abundant Corylus and presence of Taxus), whereas the abundance of Carpinus between 26.24 and 26.50 m b.s. represents Eemian pollen zone E5 (cf. Zagwijn, Reference Zagwijn1961, Reference Zagwijn and Turner1996a,Reference Zagwijn1996 b).
Between 30.40 and 30.16 m b.s., Unit 4 consists of a resistant peaty layer with wood fragments. The pollen spectra in the peat contain abundant Ulmus and Quercus pollen indicative of Eemian pollen zone E3. Interval 30.40–30.50 coincides with a reworked core-top, hence the exact boundary between the Eem Formation and Urk Formation is not preserved in the core.
Based on the combination of fine-grained sediments and diatom, pollen and malacological data, we interpret Unit 4 to mainly represent mesohaline–polyhaline estuarine or tidal-flat settings with clear water and seaweed/seagrass meadows. The lowest peaty part of the unit was formed in a swamp environment.
Unit 5 (0.00–26.24 m b.s.) (Naaldwijk Formation)
This unit is dominated by grey to light grey fine-grained to very fine-grained sand with local mollusc concentrations with abundant Peringia ulvae, Cerastoderma edule, Spisula subtruncata and Macoma balthica. Mollusc concentrations occur in intervals with coarser sand grain sizes. Tidal mud drapes are present throughout the interval. The brownish coloration in the upper c.1 m represents modern soil. The lowermost c.0.5 m of the unit is composed of organic rich brown medium-grained sand and a large displaced peat fragment which we interpret as a reworked fragment of the Early Holocene basal peat bed (Nieuwkoop Formation).
Diatoms from interval 14.00–26.25 m b.s. represent subtidal depositional settings in an area with large tidal channels (De Wolf, Reference De Wolf1996). Between 3.00 and 8.00 m b.s. a similar diatom association was found with indications for somewhat more influx of low saline waters. Pollen analysis (Veldkamp & Cleveringa, Reference Veldkamp and Cleveringa1997) shows a basal (23.15–26.13 m b.s.) Atlanticum-age succession of eutrophic/mesotrophic mire towards an oligotrophic heath. The interval 22.45–3.50 m b.s. has a Subboreal pollen assemblage with some reworked Eemian pollen, and the upper interval (1.14–3.50 m b.s.) represents high intertidal to lower supratidal environments of a Subatlanticum age.
Based on the fine-grain size, tidal mud drapes and information from diatom, pollen and malacological analysis, we interpret Unit 5 as a tidal deposit. The sharp basal contact and overlying reworked material suggest that at least the lower part of the unit was deposited in a tidal channel.
Mollusc assemblages and diversity
In total, 73 taxa at the species level (further indicated as ‘species’) were found (Supplementary Material available online at https://doi.org/10.1017/njg.2021.2). Based on NMDS, DCA and cluster analyses we found three distinct groups (Fig. 4A and B) that with the stratigraphic position in mind translate into four assemblages.
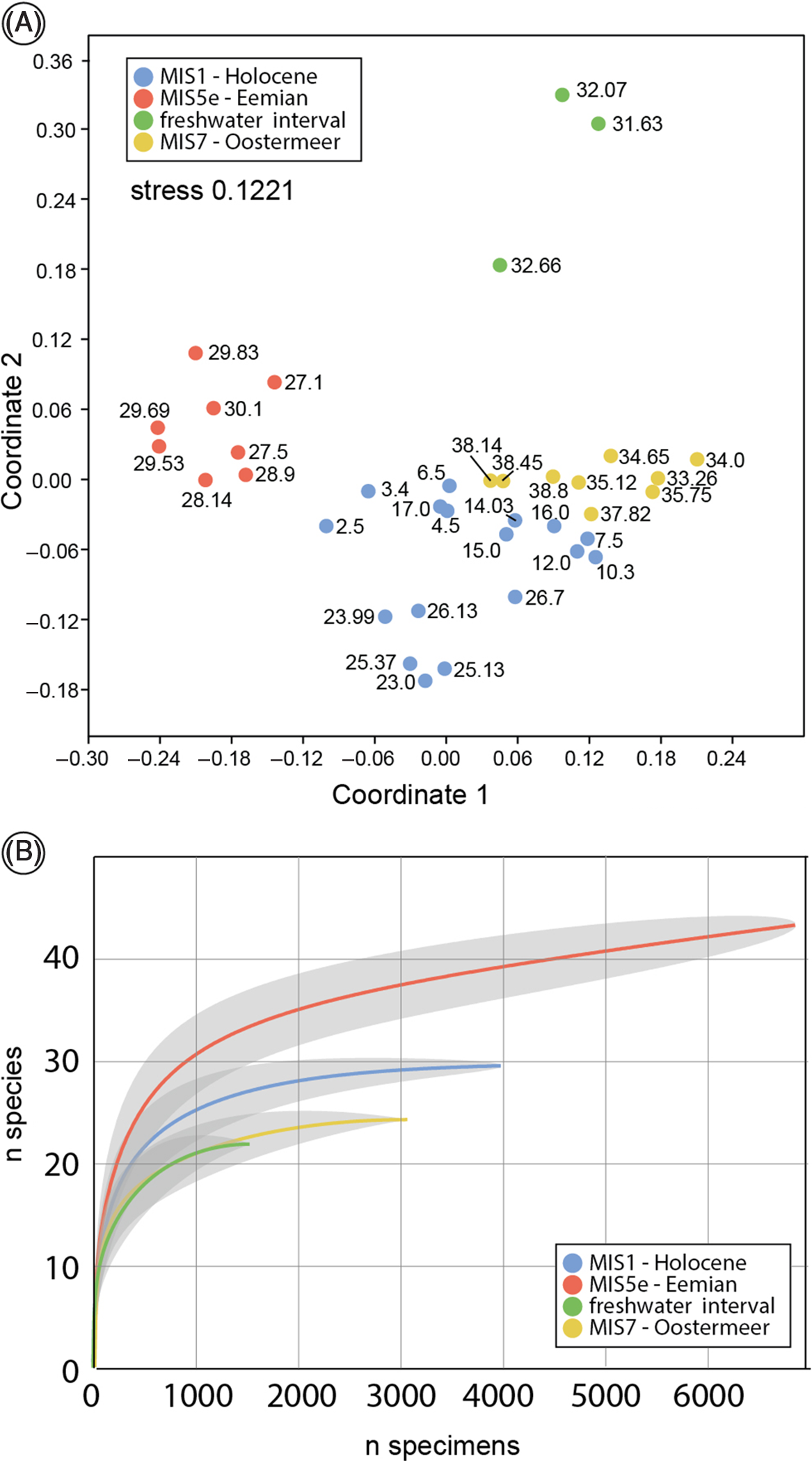
Fig. 4. (A) NMDS ordination with chord similarity measure. Numbers refer to upper depth of samples and show a clear distinction between the marine MIS5e assemblage and the overlapping MIS1/MIS7 assemblages. (B) Rarefaction analyses with 95% confidence intervals.
Assemblage 1 (2.5–25.47 m b.s.) contains 30 species, and rarefaction analysis shows that species richness appears to be saturated (Fig. 4B). It is a marine assemblage dominated by Peringia ulvae, Cerastoderma edule, Spisula subtruncata and Macoma balthica. The lower part (23–25.47 m b.s.) contains predominant mesohaline high sublittoral to low littoral species. The middle part of assemblage 1 (between 6.5 and 17.00 m) represents predominantly euhaline high sublittoral settings, and above the faunas become shallower and mesohaline. Assemblage 1 is a low boreal (Celtic) fauna type.
Assemblage 2 (26.13–30.16 m b.s.) contains 44 species, but the species richness appears not to be entirely saturated, hinting at somewhat higher richness (Fig. 4B). Dominant species are Pusillina radiate balkeii, Abra nitida, Peringia ulvae, Polititapes senescens, Cerastoderma glaucum and Bittium reticulatum s.l. Euhaline species dominate, yet polyhaline, mesohaline and oligohaline species occur. The fauna represents middle to high sublittoral conditions, and the abundance of small gastropods as well as the occurrence of Parvicardium scabrum may point to seagrass meadows and/or abundant seaweed. The fauna has a low boreal–lusitanian character, and is comparable with faunas living today in the Arcachon region, SW France (Spaink, Reference Spaink1958).
Assemblage 3 is dominated by 11 freshwater species (31.63–33.26 m b.s.) including Valvata piscinalis, five Pisidium species and several freshwater planorbiid and lymnaeiid species. The assemblage points to a lacustrine environment. Gyraulus laevis is observed to be a pioneer species from shallow ponds with a preference for stagnant electrolyte-rich (especially nitrate) freshwater (Kuijper, Reference Kuijper1973; Meijer, Reference Meijer1985). In total, 23 species occur in Assemblage 3 samples, and rarefaction analyses hints at a saturated fauna. However, some well-preserved marine species occur in this interval that are ecologically incompatible with the dominant freshwater species and must have been locally mixed in through, for example, bioturbation. The rare marine component in this fauna appears to be a subset of Assemblage 4 faunas (see below). The non-marine assemblage component is a warm-temperate Palaearctic fauna. Assemblage 3 is stratigraphically intermediate between marine MIS5e Assemblage 2 and MIS7 Assemblage 4.
Assemblage 4 (33.26–38.9 m b.s.) is a marine assemblage that contains 25 species and is dominated by Spisula subtruncata, Abra alba and Cerastoderma edule. Rarefaction suggests saturated species richness (Fig. 4B). The fauna represents mostly euhaline salinities and middle–high sublittoral conditions. It concerns a low-boreal (Celtic) assemblage.
Discussion
This study is the first full faunal analysis of a marine molluscan MIS7 assemblage from the North Sea Basin. The newly reported MIS7 fauna shows a large overlap in species and composition with MIS1 faunas: both represent similar salinity, temperature and depth regimes (MIS7 faunas representing marginally deeper conditions). Assuming that the faunas are representative for the interglacial conditions as a whole, this finding indicates that, at least on the basis of malacological evidence, MIS7 and MIS1 climate conditions were similar. The MIS7 fauna is slightly less rich in species then the MIS1 fauna. Macoma praetenuis is the sole well-preserved species occurring in MIS7 faunas that is not part of the Holocene fauna. Within the North Sea Basin, this species is well-known from the Gelasian Maassluis Formation and coeval deposits (Moerdijk et al., Reference Moerdijk, Janssen, Wesselingh, Peeters, Pouwer, van Nieulande, Janse, van der Slik, Meijer, Rijken, Cadée, Hoeksema, Doeksen, Bastemeijer, Strack, Vervoenen and ter Poorten2010; Preece et al., Reference Preece, Meijer, Penkman, Demarchi, Mayhew and Parfitt2020). The species has also been mentioned in some other apparent Middle Pleistocene localities (Meijer, Reference Meijer1993). Rare well-preserved Macoma praetenuis and similarly preserved M. obliqua from beach supplementations on the adjacent islands of Terschelling and Schiermonnikoog (de Bruyne & de Boer, Reference De Bruyne and de Boer2008; Moerdijk et al., Reference Moerdijk, Janssen, Wesselingh, Peeters, Pouwer, van Nieulande, Janse, van der Slik, Meijer, Rijken, Cadée, Hoeksema, Doeksen, Bastemeijer, Strack, Vervoenen and ter Poorten2010) likely have a Middle Pleistocene origin from faunas similar to those reported here from Ameland. Our analyses also show that MIS5e Eemian faunas are very characteristic (higher species richness, common occurrence of thermophile species) and differ strongly from MIS1 and MIS7 faunas. Our study confirms the warm-temperate character of the Eemian faunas (warmer than MIS1 and MIS7) and their use as a regional correlative assemblage.
Extending correlation over the Ameland area indicates that the Middle–Late Pleistocene succession is widely distributed in the subsurface in areas where it has not been removed by Holocene erosion (for position of tidal inlets see Vos & Knol, Reference Vos and Knol2015). The succession reported from borehole B01H0189 can be correlated with (less-detailed described) boreholes from Ameland. Eemian marine clays of the Eem Formation can be traced throughout the area and provide an important lithological marker horizon. Furthermore, the Late Saalian age sediments of the Boxtel Formation, with its well-developed coarse-grained base, and the top of the Urk Formation including the marine interval attributed here to MIS7 are lithologically recognisable and laterally traceable (Fig. 5). At one locality we even suspect the in situ presence of glacial till (Drenthe Formation, Gieten Member; Fig. 5). Our thorough reanalysis of borehole B01H0189, as well as other cores in the area, yields an interpretation that differs from the present lithostratigraphic views in the area (www.dinoloket.nl).
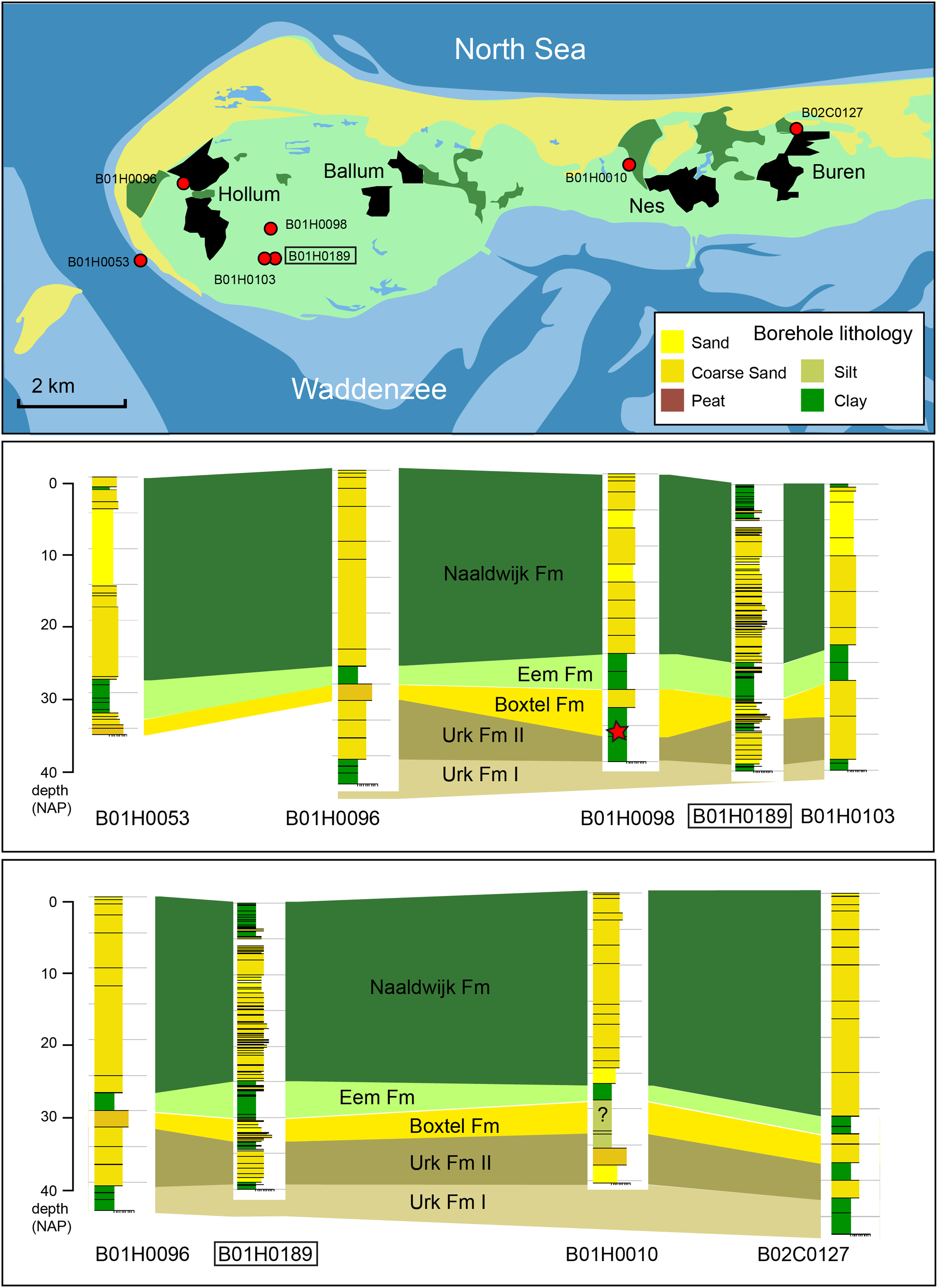
Fig. 5. Correlation of Middle Pleistocene and younger units below western Ameland. Red star denotes likely reworked Drenthe Formation gravel. All data from www.dinoloket.nl (access date 1 May 2020).
Previously, all sediments below the Holocene sequence were assigned to the Eem Formation. However, our study shows that this unit is much thinner and that at this location, most of this unit consists of Middle Pleistocene sediments of the Boxtel and Urk Formations. Implementation of our new results would improve the stratigraphic context of future mapping and modelling campaigns in the area.
In some boreholes in the region, Eemian faunas are found in a sandy interval overlying the clay interval, hence our indication of the upper boundary of the Eem Formation in older boreholes is only an approximation. The upper Pleistocene succession in many parts of the island has been eroded by Holocene tidal channels, but remains of complete stacks similar to the studied borehole indicate the presence of a MIS7–MIS5e–MIS1 succession within the subsurface of the region (Fig. 5). The Eemian fauna from borehole Hollum has many species in common with the well-studied Eemian of the central Netherlands (Cleveringa et al., Reference Cleveringa, Meijer, van Leeuwen, de Wolf, Pouwer, Lissenberg and Burger2000; Van Leeuwen et al., Reference Van Leeuwen, Beets, Bosch, Burger, Cleveringa, van Harten, Herngreen, Kruk, Langereis, Meijer, Pouwer and de Wolf2000; Wesselingh et al., Reference Wesselingh, Visser and Meijer2010). The faunas contain common thermophile species. New records for the Dutch Eemian are Akera bullata and Cylichna umbilicata.
Although direct malacological evidence for a marine connection between the North Sea and the Channel was reported by Meijer & Preece (Reference Meijer, Preece and Preece1995), malacological evidence that such a connection existed before MIS5e was absent. Although the MIS7 assemblage in itself does not directly confirm or exclude a southern marine connection, the strong resemblance to the MIS1 assemblage suggests that during MIS7 such a southern opening existed. This is in line with data from the western French Atlantic shelf (Toucanne et al., Reference Toucanne, Zaragosi, Bourillet, Gibbard, Eynaud, Giraudeau, Turon, Cremer, Cortijo, Martinez and Rossignol2009). This means that the topographic separation of the North Sea Basin and Channel region (Weald–Artois anticlinorum/Flanders–Norfolk ridge; Hijma et al., Reference Hijma, Cohen, Roebroeks, Westerhoff and Busschers2012) was already eroded deeply enough to allow marine exchange to occur. Whether or not this was preceded by fluvial downcutting during previous low stands (for which, so far, direct evidence is only known from MIS6; Busschers et al. Reference Busschers, Kasse, Van Balen, Vandenberghe, Cohen, Weerts, Wallinga, Johns, Cleveringa and Bunnik2007) or whether even older marine connections existed is the subject of further study. We need further cores (from offshore areas) to compare marine faunas from successive interglacials to assess episodic connections between the North Sea Basin and the Channel region. The occurrence of the Middle and Late Pleistocene successions in the Ameland region provides a Quaternary record that may be further explored to elucidate the palaeogeographic–climatic evolution and occupation potential of the southern North Sea Basin from the Middle to Late Pleistocene.
Conclusions
A succession of three marine intervals representing MIS7, MIS5e and MIS1 in the subsurface of Ameland (the Netherlands) enables us to compare conditions and faunas of these interglacials. The relatively widespread occurrence of the Middle and Late Pleistocene successions in the Ameland region provides a Quaternary record that sheds light on the palaeogeographic–climatic evolution of the southern North Sea Basin. The faunas of MIS7 are very similar to those of MIS1; MIS5e is more diverse and represents warmer conditions. The stratigraphic attribution of a warm-temperate freshwater fauna that occurs in between the MIS7 and MIS5e intervals is not yet fully resolved, but most likely represents terminal MIS6 intervals. No direct faunal evidence of an MIS7 connection through the Channel region to the south exists, but the resemblance to Holocene faunas makes this connection distinctly possible.
Acknowledgements
We thank the editor and two anonymous reviewers who provided us with excellent suggestions that helped to improve the manuscript.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/njg.2021.2








 Open access
Open access