



Introduction
Soybean [Glycine max (L.) Merr.] production provides a base for global food security because it is the most important source of vegetable protein, widely used in food and feed products (Beta and Isaak, Reference Beta, Isaak, Wrigley, Corke, Seetharaman and Faubion2016; Smárason et al., Reference Smárason, Alriksson and Jóhannsson2019; Parisi et al., Reference Parisi, Tulli, Fortina, Marino, Bani, Dalle Zotte, Angelis, Piccolo, Pinotti, Schiavone, Terova, Prandini, Gasco, Roncarati and Danieli2020; Wajid et al., Reference Wajid, Ahmad, İftakhar, Qureshi and Ceyhan2020). Brazil is the largest soybean producer in the world, with over 41.5 million ha (based on the 2021/2022 harvest) in tropical or subtropical environments, it accounts for 36% of global soybean production (USDA, 2021). It is estimated that the world's population will rise from 7.7 to 9.8 billion between 2017 and 2050 (United Nations, 2017); this will require a considerable increase in the quantity and quality (protein and oil concentration) of food production (United Nations, 2017). In this sense, with limited new land on which to expand and an emphasis on sustainable systems, increases in soybean production should come primarily from increased production per unit area, requiring sustainable agricultural intensification (Cassman and Grassini, Reference Cassman and Grassini2020; Marin et al., Reference Marin, Zanon, Monzon, Andrade, Figueiredo Moura da Silva, Richter, Antolin, Ribeiro, Ribas, Battisti, Heinemann and Grassini2022).
Seed protein and oil concentration play a significant role in determining the quality of soybean food products (Clarke and Wiseman, Reference Clarke and Wiseman2000; Friedman and Brandon, Reference Friedman and Brandon2001; Li et al., Reference Li, Gu, Wu, Qin, Guo, Wang, Zhang and Jiang2021). Usually, the soybean quality is measured by the amount of oil extracted and final crude protein concentrations in the soybean meal (Singh and Koksel, Reference Singh and Koksel2021; Wen et al., Reference Wen, Liu, Meng, Li and He2021). Some soybean importers currently offer premiums for soybean containing higher amounts of protein (Pathan et al., Reference Pathan, Vuong, Clark, Lee, Shannon, Roberts, Ellersieck, Cregan, Hyten, Nguyen and Sleper2013; William et al., Reference William, Dahl and Hertsgaard2020; Lakkakula et al., Reference Lakkakula, Bullock and Wilson2021), while some countries exclude or apply penalties to soybeans from regions when quality requirements are not met. China, for instance, requires minimums of 33.5% of protein and 18% of oil concentration (Hertsgaard et al., Reference Hertsgaard, Wilson and Dahl2019). Therefore, advances in our understanding of the effects of agricultural practices on soybean seed quality under tropical and subtropical environments can contribute to food security and agribusiness.
Soybean seed yield is linearly related to the total plant N uptake (Salvagiotti et al., Reference Salvagiotti, Cassman, Specht, Walters, Weiss and Dobermann2008; Tamagno et al., Reference Tamagno, Balboa, Assefa, Kovács, Casteel, Salvagiotti, García, Stewart and Ciampitti2017). The soybean crop obtains N from the symbiotic fixation of atmospheric N2 (SNF), soil N mineralization and N from irrigation or water table; not usually relying upon N-fertilizer application (Herridge et al., Reference Herridge, Peoples and Boddey2008; Randall, et al., Reference Randall, Delgado and Schepers2008; Reis et al., Reference Reis, Rosso, Purcell, Naeve, Casteel, Kovacs, Archontoulis, Davidson and Ciampitti2021). Soybeans derive between 25 and 75% of their total nitrogen from SNF process (Deibert et al., Reference Deibert, Bijeriego and Olson1979; Salvagiotti et al., Reference Salvagiotti, Cassman, Specht, Walters, Weiss and Dobermann2008; Collino et al., Reference Collino, Salvagiotti, Perticari, Piccinetti, Ovando, Urquiaga and Racca2015). However, there has been a concern recently about whether SNF would be sufficient to meet the increased N needs for high-yielding soybean crops (Cafaro La Menza et al., Reference Cafaro La Menza, Monzon, Specht and Grassini2017, Reference Cafaro La Menza, Monzon, Specht, Lindquist, Arkebauer, Graef and Grassini2019, Reference Cafaro La Menza, Monzon, Lindquist, Arkebauer, Knops, Unkovich, Specht and Grassini2020; Basal and Szabó, Reference Basal and Szabó2020). These studies were conducted under temperate environments and reported that N-fertilization could contribute to increasing soybean yield. Temperate soils are generally viewed as more favourable to crop production than tropical because of higher nutrient levels (Rosenzweig and Liverman, Reference Rosenzweig and Liverman1992; Caubet et al., Reference Caubet, Cornu, Saby and Meunier2020). In contrast, soils in humid tropics tend to be highly leached of nutrients due to high temperatures, intense rainfall and erosion that accelerate the decay of soil organic matter, resulting in a decreased C:N ratio (Rosenzweig and Liverman, Reference Rosenzweig and Liverman1992; Alemayehu et al., Reference Alemayehu, Zerihun and Solomon2018). The N-fertilization on tropical soybean has been little explored in scientific papers.
The consistent increase in soybean yield and decrease in seed protein concentration often raise a debate on the need for extra N supply. In temperate environments, with maturity group (MG) ranging from I to IV, N-fertilization on soybean has been widely explored. For instance, Ray et al. (Reference Ray, Heatherly and Fritschi2006) and Salvagiotti et al. (Reference Salvagiotti, Specht, Cassman, Walters, Weiss and Dobermann2009) obtained seed yield increase between 130 and 438 kg/ha, while Cafaro La Menza et al. (Reference Cafaro La Menza, Monzon, Specht and Grassini2017, Reference Cafaro La Menza, Monzon, Specht, Lindquist, Arkebauer, Graef and Grassini2019) obtained seed yield increase around 600 kg/ha, and an average seed protein concentration increase of 0.9 g/100 g with N-fertilizer and under irrigated conditions. Further, Córdova et al. (Reference Córdova, Archontoulis and Licht2020) obtained soybean yield increase ranging from 300 to 1000 kg/ha, while in a recently published study, Pannecoucque et al. (Reference Pannecoucque, Goormachtigh, Ceusters, Bodé, Boeckx and Roldán-Ruiz2022) obtained an average increase in seed yield of 209 kg/ha and a decrease of 2.0 g/100 g of protein concentration, using seed inoculation and 35 kg N/ha at R1. Finally, Wesley et al. (Reference Wesley, Lamond, Martin and Duncan1998) and Brar and Lawley (Reference Brar and Lawley2020) reported non-significant N effects on seed yield and protein concentration. In tropical and subtropical environments (with MG ranging from V to VIII), efforts have been made by a few researchers to understand the soybean responses to N-fertilization. Cordeiro and Echer (Reference Cordeiro and Echer2019) obtained 439 kg/ha increase in soybean yield (there is no information about protein concentration) when N-fertilization was combined with more efficient N-fixing bacteria rates under unfavourable environments, while Hungria et al. (Reference Hungria, Franchini, Campo, Crispino, Moraes, Sibaldelli, Mendes and Arihara2006) and Zilli et al. (Reference Zilli, Pacheco, Gianluppi, Smiderle, Urquiaga and Hungria2021) applied 200 kg N/ha and did not obtain any seed yield increase (there is no information about protein concentration).
The N limitation for soybean in temperate environments is driven by the yield level (larger limitation in high than lower yield levels) and indigenous soil N supply of the production environment (Cafaro La Menza et al., Reference Cafaro La Menza, Monzon, Specht and Grassini2017, Reference Cafaro La Menza, Monzon, Specht, Lindquist, Arkebauer, Graef and Grassini2019). Still, it is unclear if these drivers also apply to tropical and subtropical environments. Seed protein concentration reports in soybean studies with N-fertilization under tropical and subtropical environments are scarce, and there is comparatively less research (and without a consistent protocol) on soybean seed yield responses to N supply in tropical and subtropical than in temperate environments. Our scientific question is how N supply could improve soybean yields and protein concentration in tropical and subtropical conditions. Therefore, this research adopted consistent N-fertilization treatments applied to soybean grown in tropical and subtropical environments under irrigation or rainfed condition in Brazil to investigate whether N supply can significantly increase seed yield and/or seed protein or oil concentration (latitude range: 12°S to 29°S). We sought to test the hypothesis that soybean seed yield and/or protein concentration are limited by N supply under tropical and subtropical conditions. Our goal was to evaluate the effects of N supply on soybean yield, and seed quality under rainfed and irrigated conditions and across 17 degrees of latitude range of tropical and subtropical environments in Brazil.
Materials and methods
Field experiments
We conducted 11 field experiments under rainfed or irrigated conditions with a two-way treatment (environment × N-fertilization) structure in a randomized complete block design with four replicates. The experimental plot consisted of 12 rows, 9.0 m long, with a spacing of 0.5 m between rows and 0.04 m for planting depth. The environments were defined as the combination of locations, seasons, sowing dates and cultivars; with different crop management:
(i) Sorriso (12°42′S, 55°48′W, 375 a.m.s.l), during two rainfed cropping seasons [2018/2019 (SO-1) and 2019/2020 (SO-2)], with climate classification of Aw (tropical savanna climate), conducted under conventional tillage on Dystrophic Red Yellow Ferrosol, where the crop before planting was an 8-year degraded pasture area for SO-1, and maize for SO-2;
(ii) Piracicaba (22°42′S, 47°30′W, 546 a.m.s.l), during three rainfed and three irrigated cropping seasons [2017/2018 (PI-1), 2018/2019 (PI-2) and 2019/2020 (PI-3)] with a climate classification of Cwa (high-altitude tropical) (Koeppen, 1948), conducted under conventional tillage on a Eutric Rhodic Ferralic Nitisol, the previous crop was maize (Zea mays L.) for P1-1 and PI-3, and wheat (Triticum aestivum L.) for PI-2, area with a 5-year history of soybean yield >4.5 Mg/ha;
(iii) Cruz Alta (28°38′S, 53°36′W, 476 a.m.s.l), during one rainfed cropping season [2018/2019 (CA-1)], with a climate classification of Cfa (humid subtropical, with well-defined summer and winter seasons) (Koeppen, 1948), conducted under no-tillage practices on Dystrophic Red Acrisol, the previous crop was wheat, area with a 28-year history of soybean yield >4.5 Mg/ha;
(iv) Tupanciretã (28°48′S, 53°48′W, 466 a.m.s.l) during one rainfed and one irrigated cropping season [2019/2020 (TU-1)], with a climate classification of Cfa, conducted under no-tillage on Dystrophic Red, the previous crop was maize, area with a 26-year history of soybean yield >4.5 Mg/ha (Table 1 and Fig. 1).
Fig. 1. Soybean harvest area distribution in Brazil. Data source: Brazilian Institute of Geography and Statistics (Instituto Brasileiro de Geografia e Estatística, 2021). Black stars indicate field experiments.
Table 1. Description of field experiments conducted in Piracicaba, Sorriso, Cruz Alta and Tupanciretã

MG, maturity group.
a Time between VE (emergence) and R7 (beginning of maturity), using phenological stages proposed by Fehr and Caviness (Reference Fehr and Caviness1977).
The irrigation amounts applied in the experiments were determined by the potential evapotranspiration determined by the Priestley and Taylor (Reference Priestley and Taylor1972) method; computed using the daily weather data measured with well-calibrated sensors from the on-site weather station (see Table 1 and Supplementary Figs S1–S5). The Priestley–Taylor method was applied under minimum advection conditions and using empirical parameter α = 1.26 (Pereira and Villa Nova, Reference Pereira and Villa Nova1992; Figueiredo Moura da Silva et al., Reference Figueiredo Moura da Silva, Gonçalves, Pereira, Fattori Júnior, Sobenko and Marin2019, Reference Figueiredo Moura da Silva, Boote, Hoogenboom, Gonçalves, Junior and Marin2021, Reference Figueiredo Moura da Silva, Hoogenboom, Boote, Gonçalves and Marin2022). The water amount required was applied with sprinkler irrigation by centre pivot sprinklers Senninger Model i-Wob-UP3 at Piracicaba, and centre pivot sprinklers Plona KS 1500 at Tupanciretã.
N-fertilizer treatments were defined as (i) no N-fertilizer (0N), where the soybean only relied on indigenous soil N sources (inorganic soil N, mineralized N and SNF); and (ii) with N-fertilizer (1000N). The total amount of 1000 kg N/ha fertilizer was calculated using a soybean yield target of 8945 kg/ha; this value was based on the maximum soybean yield documented in Brazil, until the season 2016/2017, in contest areas in Brazil (CESB, 2017 and later reported by Battisti et al., Reference Battisti, Sentelhas, Pascoalino, Sako, de Sá Dantas and Moraes2018). The N-fertilizer source was urea (46-0-0) applied on the surface. It is noteworthy that our objective was not to replicate the management of soybean contest areas, we only sought a maximum yield seed reference to compute supplementing N-fertilizer on soybeans.
We assumed that the crop accumulates 79 kg N/ha in its aboveground biomass per each additional 1000 kg of seed yield (Salvagiotti et al., Reference Salvagiotti, Cassman, Specht, Walters, Weiss and Dobermann2008; Tamagno et al., Reference Tamagno, Balboa, Assefa, Kovács, Casteel, Salvagiotti, García, Stewart and Ciampitti2017), with an extra 40% of N applied to compensate for N losses through ammonia volatilization (Cantarella et al., Reference Cantarella, Otto, Soares and de Brito Silva2018), and another 1.5% N applied to compensate N losses by leaching and runoff (Libardi and Reichardt, Reference Libardi and Reichardt1978; Oliveira et al., Reference Oliveira, Trivelin and Oliveira2007). The urea amount applied was split into five applications during the cropping season following crop N demand reported by Thies et al. (Reference Thies, Singleton and Bohlool1995) and Bender et al. (Reference Bender, Haegele and Below2015): (i) 10% in second-node (V2); (ii) 10% in fourth-node (V4); (iii) 20% in full bloom stage (R2); (iv) 30% in beginning pod stage (R3); (v) 30% in beginning seed stage (R5). For both treatments, the seeds were inoculated with Bradyrhizobium elkanii (strains SEMIA 587 and SEMIA 5019) at a concentration of 5 × 109 CFU/ml (colony-forming units).
Macronutrient fertilization was computed based on surface and subsurface soil analysis. Soil nutrient initial levels among all experiments were as follows: 0.9–3.1% of organic matter (Walkey–Black method), 0.7–27.0 mg P/dm3 (Melich-1 method), 0.2–5.4 K mmolc/dm3 (Melich-1 method), 4.0–19 Mg mmolc/dm3 (extracted by calcium acetate), 10.6–44.2 Ca mmolc/dm3 (extracted by calcium acetate), 3.0–15.0 mg S/kg (turbidimetry method) (see Supplementary Table S1). The soil fertilization sought to reach: P > 26 mg/m3, K > 2.5 mmolc/dm3, Mg > 13 mmol/dm3, Ca > 26 mmolc/dm3, S > 15 mmol/dm3. The soybean micronutrients were supplied via foliar fertilization based on the crop cycle and extraction and export of micronutrients (see Supplementary Table S2).
Water deficit was computed as the difference between water input (precipitation and/or irrigation) and computed potential evapotranspiration. Phenological stages, according to Fehr and Caviness (Reference Fehr and Caviness1977), were tracked every 5 days in all experiments and recorded dates of emergence (VE), beginning flowering (R1), beginning pod (R3), beginning seed (R5) and physiological maturity (R7) (Fig. 2), when 50% of the plants were at each stage (Fehr and Caviness, Reference Fehr and Caviness1977). Field measurements for each experiment included (i) seed yield (kg/ha), (ii) seed protein concentration (Kjeldahl method) and (iii) seed oil concentration (Goldfish method). The seed yield, seed protein and oil concentration values were expressed at 0.13 kg H2O/kg seed. Seed yield, seed protein and oil concentration samples were collected (hand-harvested) on three centre rows (9 m2) of the experimental plot for each treatment and replication of each season.
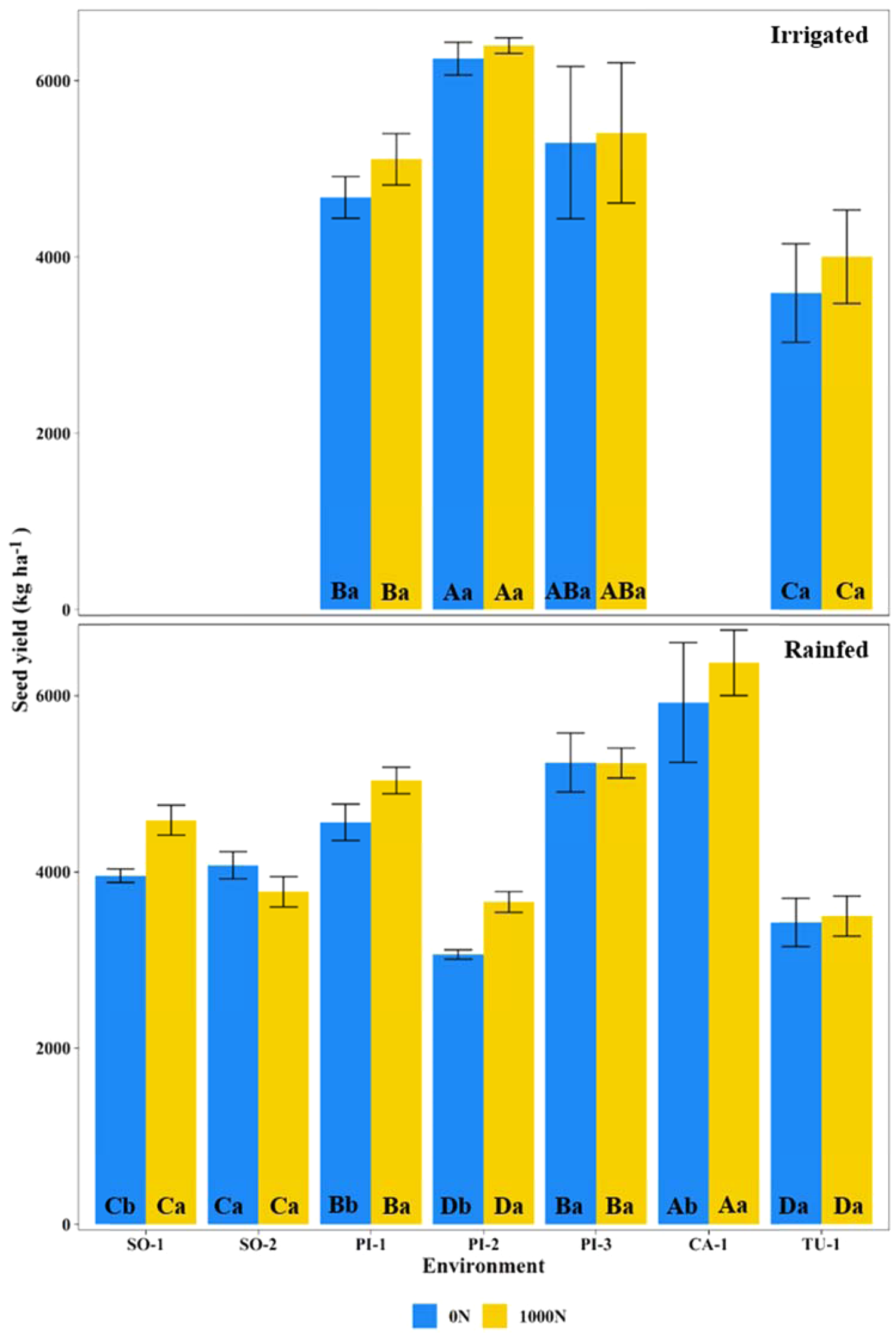
Fig. 2. Seed yield under irrigated or rainfed conditions, with zero (0N) or 1000 kg N/ha (1000N). For experiments conducted in Piracicaba (PI) under three seasons, Sorriso (SO) under two seasons, Cruz Alta (CA) under one season, and Tupanciretã (TU) under one season. The experiments were shown from the locations of low latitude (left) to high latitude (right). Means followed by different letters, uppercase among environments and lowercase between N-treatment for each environment, differ by Tukey test (P < 0.05).
Statistical analysis
Bartlett test (Milliken and Johnson, Reference Milliken and Johnson2009) was applied to investigate the homogeneity of variances, and no significant differences were observed (P > 0.05) for all experiments. Once the homogeneity of within-environment variances was obtained, we conducted the combined analysis of variance to determine the effect of environments and N treatments on seed yield, seed protein and oil concentration; and the means were compared by the Tukey test considering the significance level of 5% (agricolae package, Team R Core, https://www.r-project.org). The N-fertilizer and environment were considered a fixed effect, and blocks were considered a random effect. Linear regression was used to estimate the relation: (i) between seed protein or oil concentration and water deficit; and (ii) between seed protein or oil concentration and latitude.
Results
The higher N supply in the 1000N treatment significantly increased the seed yield by 277 kg/ha (7%) and the seed protein concentration by 4.3 g/100 g (12%) in tropical and subtropical environments in Brazil under rainfed conditions (Table 2, Figs 2 and 3). However, the magnitude of the response varied according to the environment from −300 kg/ha (−7%) to 596 kg/ha (+20%) for seed yield and from 2.2 g/100 g (6%) to 6.2 g/100 g (16%) for protein. In the case of irrigated experiments, the increased N supply in the 1000N treatment did not affect the seed yield but increased seed protein concentration by 1.6 g/100 g (4%) (Table 2, Figs 2 and 3). Furthermore, the magnitude of responses in seed oil concentration due to N supply also varied across irrigated and rainfed conditions from −1.2 g/100 g (−8%) to 1.8 g/100 g (+11%). While seed yield responses to 1000N were significant (in some environments under rainfed) or not significant (in irrigated), the seed protein concentration consistently increased across both water availability conditions.

Fig. 3. Seed protein concentration under irrigated or rainfed conditions, with zero (0N) or 1000 kg N/ha (1000N). For experiments conducted in Piracicaba (PI) under three seasons, Sorriso (SO) under two seasons, Cruz Alta (CA) under one season and Tupanciretã (TU) under one season. The experiments were shown from the locations of low latitude (left) to high latitude (right). Means followed by different letters, uppercase among environments and lowercase between N-treatment for each environment, differ by Tukey test (P < 0.05).
Table 2. Analysis of variance for effects of environments, nitrogen (N)-fertilization treatment and interaction between environments × N-fertilization in randomized complete block design structure in experiments with rainfed conditions

d.f., degrees of freedom; F, F-statistic; MS, mean square.
Statistical significance was indicated by: *P < 0.05; **P < 0.01; ***P < 0.001.
We observed that 1000N treatment only promoted a significant seed yield increase at 476 kg/ha for PI-1, 596 kg/ha for PI-2, 627 kg/ha for SO-1 and 452 kg/ha for CA-1 (Fig. 2). The increase in N supply through the 1000N treatment provided statistically significant gains in seed protein concentration under irrigated and rainfed conditions (P < 0.001; Table 2; Fig. 3). The average seed protein concentration was 41.0 g/100 g for 1000N and 39.4 g/100 g for 0N treatment under irrigated condition and 40.5 g/100 g for 1000N and 36.2 g/100 g for 0N under rainfed conditions. The average seed protein concentration increased in the 1000N compared to the 0N treatment on 1.6 g/100 g in irrigated experiments and 4.3 g/100 g in rainfed experiments. However, the magnitude of the treatment response varied from 2.2 to 6.5 g/100 g in rainfed experiments.
For experiments conducted under irrigated environments, the seed oil concentration ranged from 14.1 to 18.7 g/100 g (Fig. 4). The N treatments provided no statistically significant results for seed oil concentration under irrigated conditions. Under rainfed conditions, seed oil concentrations ranged from 14.8 to 20.0 g/100 g (Fig. 4). The response to N treatments on seed oil concentration was unclear. The average seed oil concentration obtained in rainfed environments was 17.8% with 0N and 17.7% with 1000N.
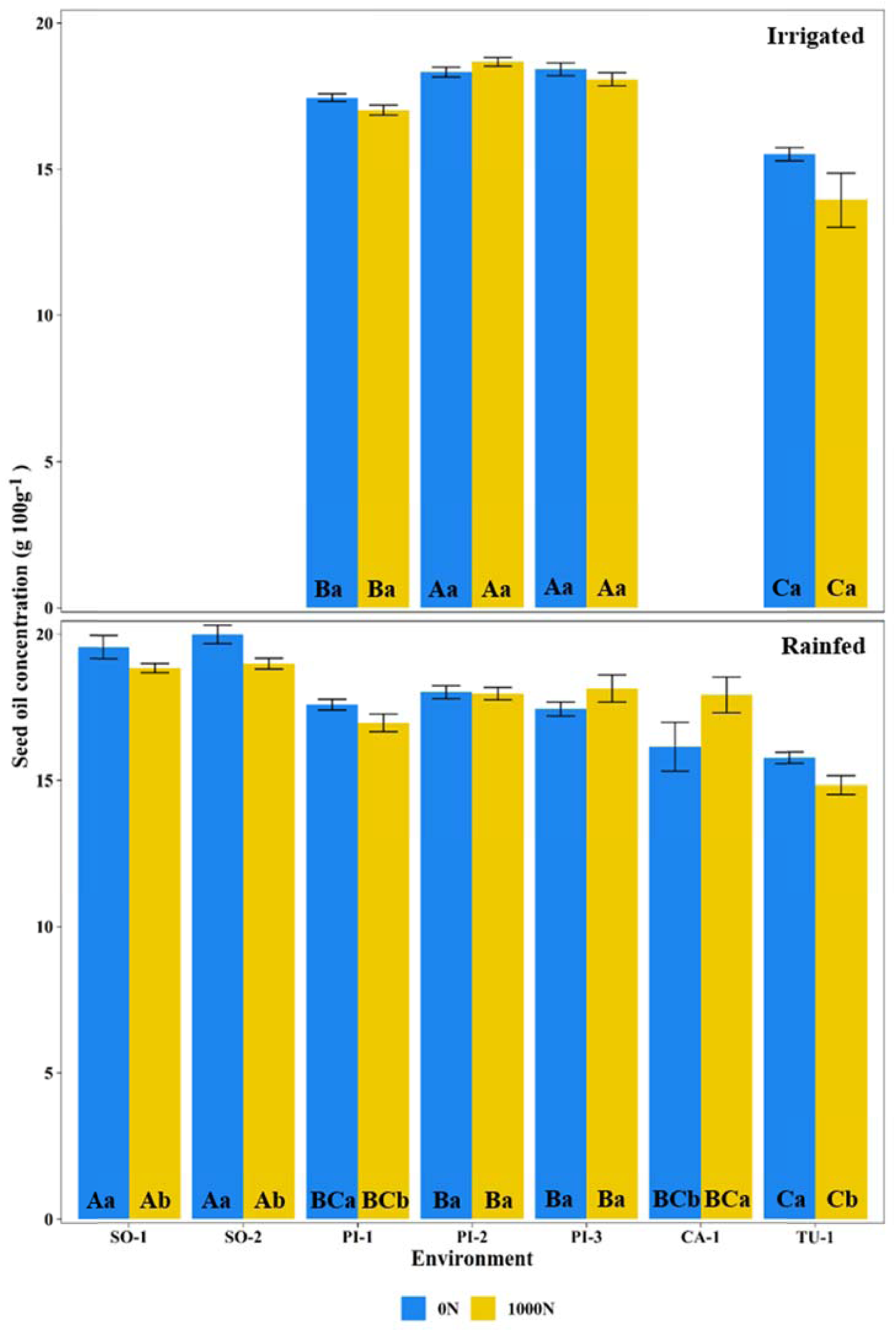
Fig. 4. Seed oil concentration under irrigated or rainfed conditions, with zero (0N) or 1000 kg N/ha (1000N). For experiments conducted in Piracicaba (PI) under three seasons, Sorriso (SO) under two seasons, Cruz Alta (CA) under one season and Tupanciretã (TU) under one season. The experiments were shown from the locations of low latitude (left) to high latitude (right). Means followed by different letters, uppercase among environments and lowercase between N-treatment for each environment, differ by Tukey test (P < 0.05).
There was a positive linear relationship between the seed protein concentration difference ((1000N − 0N)/0N × 100) and the water deficit, with R 2 = 0.84 (Fig. 5). The relative response in seed protein concentration due to N supply increased by 4% for every 100 mm of water deficit. For seed oil concentration, the water deficit did not explain the variability.

Fig. 5. Variation (Δ) of seed protein concentration (%) between 1000N and 0N treatments (1000N minus 0N) with water deficit across all rainfed conditions (close circles) in this study (total of seven environments). The Δ of seed protein concentration in irrigated environments is also shown (open circles). Water deficit was calculated as the difference between water input (precipitation and/or irrigation) and potential evapotranspiration using Priestley–Taylor method. A linear regression adjusted to the rainfed environments indicates the increase in Δ of seed protein concentration due to N supply with increasing water deficit.
We also found that a large portion of the variability in seed protein and oil concentration across environments under rainfed or irrigated conditions was related to the latitude in which the experiments were conducted (Figs 6(a) and (b)). It is important to note that the relationship was established using only four observations representing the sites. The higher seed protein and oil concentration values occurred in lower latitudes (12°S). The soybean harvested across Southern Brazil experiments showed consistently lower seed protein and oil concentrations than Northern Brazil. The magnitude of the decrease in seed protein concentration with latitude was slightly higher than in oil (−0.34× v. −0.20×).
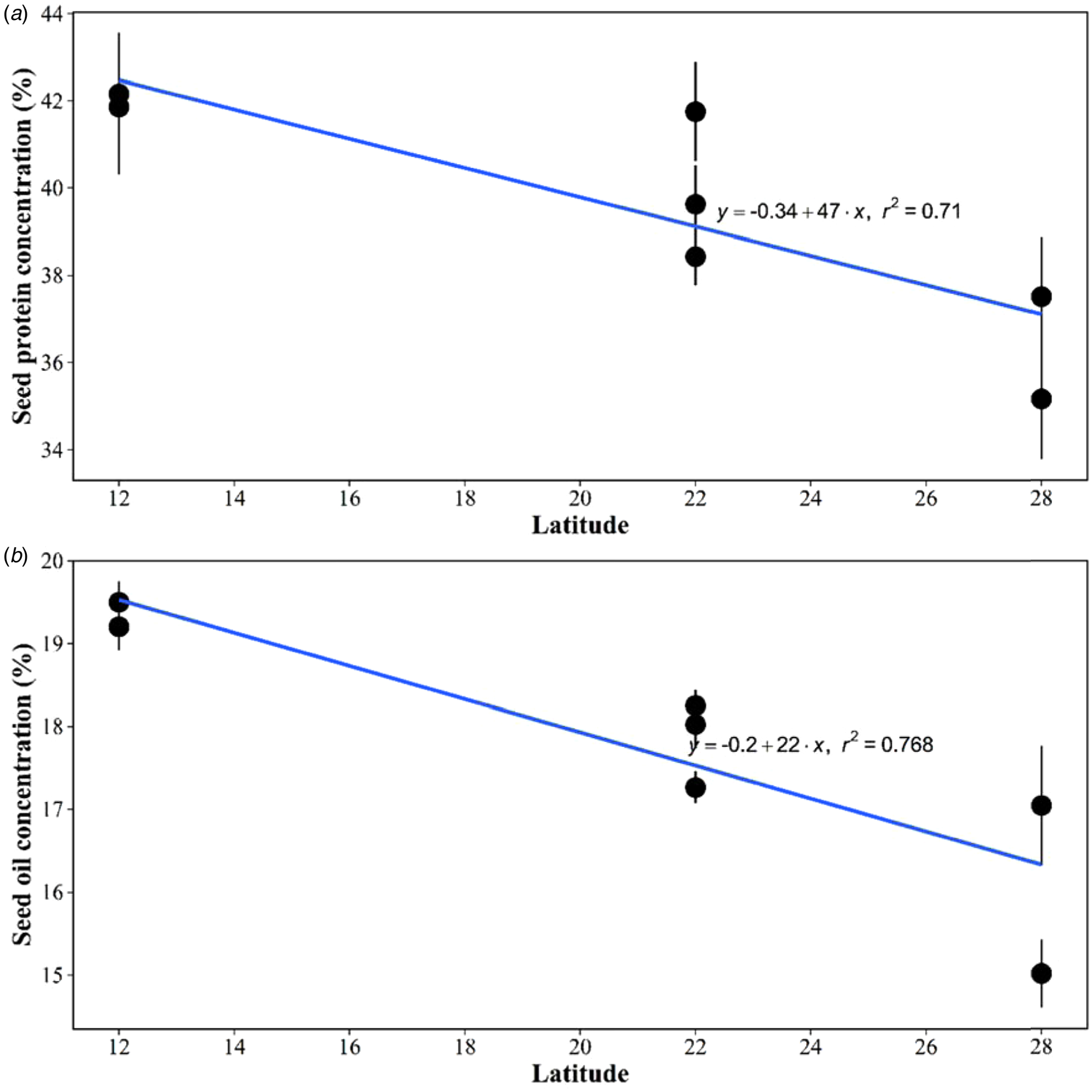
Fig. 6. Average of seed protein (a) and oil concentration (b) (g/100 g) for experiments conducted under rainfed or irrigated conditions across different latitudes. Each average includes the four replicates of the 0N and 1000N treatments. Linear regression was adjusted to each relationship between seed protein and oil to latitude.
Discussion
Unlike in temperate environments, soybean seed yield failed to respond to increases in N supply in most tropical and subtropical environments evaluated in this study. Salvagiotti et al. (Reference Salvagiotti, Specht, Cassman, Walters, Weiss and Dobermann2009) and Cafaro La Menza et al. (Reference Cafaro La Menza, Monzon, Specht and Grassini2017, Reference Cafaro La Menza, Monzon, Specht, Lindquist, Arkebauer, Graef and Grassini2019, Reference Cafaro La Menza, Monzon, Lindquist, Arkebauer, Knops, Unkovich, Specht and Grassini2020) documented the existence of N limitation in highly productive environments in trials in Argentina and the USA, with N-fertilization under irrigated conditions. Their findings revealed the potentiality of soybean seed yield increases with ample N supply in temperate environments with a yield level of >4.5 Mg/ha (based on the yield history of the area). In our areas with yield level >4.5 Mg/ha, the seed yield responded to the 1000N treatment only in PI-3 (rainfed conditions) and Cruz Alta. In these sites, we sowed cultivars with MGs between 6.2 and 6.5, which are much longer MGs than the ones used in previous studies in temperate zones (MGs between 0.0 and 3.8). For PI-1, PI-2 and TU-1, the indigenous soil N sources were sufficient to reach the same seed yield level as soybeans with N-fertilizer. Still, the yield target of 8945 kg/ha was not reached in any experiment. It means that the sowing date may have been a limiting factor in reaching higher yield; because for the Southern Hemisphere, sowing between late September and mid-October may be more ideal for the soybean crop to reach its peak of development (between R5 and R6) in December (greatest solar energy availability) (Zanon et al., Reference Zanon, Streck and Grassini2016; Tagliapietra et al., Reference Tagliapietra, Zanon, Streck, Balest, Rosa, Bexaira, Richter, Ribas and Silva2021). Therefore, soybean could have accumulated more carbohydrates and acquire N via fixation or fertilization.
In our experiments, the SNF could have been indirectly affected in the few cases where increased seed protein also accompanied seed yield response to N-fertilization, for instance in SO-1. This environment was limited for SNF for two reasons: firstly, although the seeds were inoculated with B. elkanii, the soil probably had a poor inoculum of N-fixing bacteria because, before planting, it was a degraded pasture area with no prior soybean crop history (Cordeiro and Echer, Reference Cordeiro and Echer2019). Secondly, a low initial soil phosphorus content (see Supplementary Table S1) may reduce root nodulation (Hungria et al., Reference Hungria, Franchini, Campo, Crispino, Moraes, Sibaldelli, Mendes and Arihara2006; Pavanelli and Araújo, Reference Pavanelli and Araújo2009). Although the soil was fertilized with phosphorus, it cannot be certain that the element was available in the early stages of inoculation. Consistent with our results, Cordeiro and Echer (Reference Cordeiro and Echer2019) obtained an increase of 439 kg/ha (22%) in the soybean yield using N-fertilizer (50 kg/ha) in the first year of sowing in areas following degraded pasture.
During the cropping season at PI-2, there were water-deficit periods between R3 and R6 [critical period for N accumulation and seed number determination (Rotundo et al., Reference Rotundo, Borrás, De Bruin and Pedersen2012; Monzon et al., Reference Monzon, La Menza, Cerrudo, Canepa, Edreira, Specht, Andrade and Grassini2021; Egli, Reference Egli2017)], the accumulated rainfall was only 54.3 mm, with 97.8% of rainfall concentration on 77 and 84 DAP (Fig. 2; Supplementary Fig. S1-b). Water stress affects the survival and synthesis of leghaemoglobin and nodule function (Sprent, Reference Sprent1971; Patterson and Hudak, Reference Patterson, Hudak, Elkan and Upchurch1996; Santachiara et al., Reference Santachiara, Salvagiotti and Rotundo2019). Similarly, several studies have shown that N uptake and assimilation from the soil are less sensitive to temporary water deficits than SNF (Purcell and King, Reference Purcell and King1996; Purcell et al., Reference Purcell, Serraj, Sinclair and De2004; Ray et al., Reference Ray, Heatherly and Fritschi2006). However, it is still unclear how much yield loss can be consistently captured with an economically feasible N-fertilizer amount. Also unknown, is which type, timing and placement of the N-fertilizer would maximize yield response and minimize N losses in rainfed environments that usually experience diverse drought conditions (amount and timing of water deficit).
N-fertilization on soybean could impact seed composition despite not necessarily affecting seed yields (Assefa et al., Reference Assefa, Purcell, Salmeron, Naeve, Casteel, Kovács, Archontoulis, Licht, Below, Kandel, Lindsey, Gaska, Conley, Shapiro, Orlowski, Golden, Kaur, Sigh, Thelen, Laurenz, Davidson and Ciampitti2019). We found an average potential increase of 1.6 g/100 g (4%) in soybean seed protein concentration due to N-fertilizer application under experiments conducted under irrigated conditions. This potential increase was ca. 60% higher than the average of 0.9 g/100 g documented by Cafaro La Menza et al. (Reference Cafaro La Menza, Monzon, Specht and Grassini2017, Reference Cafaro La Menza, Monzon, Specht, Lindquist, Arkebauer, Graef and Grassini2019, Reference Cafaro La Menza, Monzon, Lindquist, Arkebauer, Knops, Unkovich, Specht and Grassini2020) with N-fertilization under irrigated conditions in temperate environments. Yet, in our experiments conducted under rainfed conditions, we found a larger potential response to N-fertilizer in terms of seed protein concentration (4.3 g/100 g). This potential response was twofold higher than the 2.0 g/100 g documented by Bosaz et al. (Reference Bosaz, Borrás, Gerde, Santachiara and Rotundo2021) under rainfed conditions in a temperate environment.
Different from results obtained for seed protein concentration, our findings showed unclear relation between N supply and seed oil concentration: (1) no response in all experiments under irrigated conditions and PI-2 under rainfed conditions, (2) negative response in SO-1, SO-2, PI-1 and TU-1 under rainfed conditions, and (3) positive response in PI-3 and CA-1 under rainfed conditions. In a meta-analysis study, Rotundo and Westgate (Reference Rotundo and Westgate2009) reported that water stress decreased seed oil concentration dramatically for soybean grown in the USA. However, their results were not consistent with ours in subtropical and tropical environments in which the water deficit did not consistently affect seed oil concentration. The seed oil and protein concentration can be affected by genotype, environment, management and season in a complex and still unclear way (Rotundo and Westgate, Reference Rotundo and Westgate2009; Rotundo et al., Reference Rotundo, Miller Garvin and Naeve2016; Assefa et al., Reference Assefa, Purcell, Salmeron, Naeve, Casteel, Kovács, Archontoulis, Licht, Below, Kandel, Lindsey, Gaska, Conley, Shapiro, Orlowski, Golden, Kaur, Sigh, Thelen, Laurenz, Davidson and Ciampitti2019; Bosaz et al., Reference Bosaz, Borrás, Gerde, Santachiara and Rotundo2021; Grassini et al., Reference Grassini, Cafaro La Menza, Rattalino Edreira, Monzón, Tenorio, Specht, Sandras and Calderini2021). Unlike protein, the interaction between N supply, latitudes and water deficit for seed oil did not follow a clear pattern in tropical and subtropical environments.
Our findings highlighted that N supply is a critical factor in tropical and subtropical soybean systems to increase the seed protein concentration and that the protein gap due to N supply is much larger in tropical and subtropical than in temperate environments. Indeed, more research efforts are needed to diagnose nutrient limitations in tropical and subtropical environments and test N-fertilization timing, rates, placement and fertilizer types for an economical and environmentally sound response (Rotundo et al., Reference Rotundo, Rech, Cardoso, Fang, Tang, Olson, Pyrik, Conrad, Borras, Mihura and Messina2022). At the cropping system level, increasing indigenous soil N supply with cover crops or different crop rotations and developing novel bacteria strains with improved SNF performance and survival in the soil will be needed to close the N gap for seed yield and protein in soybean.
Yet, our findings also revealed that the soybean harvested in lower latitudes showed consistently higher seed protein and oil concentrations than in higher latitudes within tropical and subtropical environments. It should be noted that we used only four points to establish this relationship, so this trend needs to be confirmed with more data in tropical conditions. In a meta-analysis, Assefa et al. (Reference Assefa, Purcell, Salmeron, Naeve, Casteel, Kovács, Archontoulis, Licht, Below, Kandel, Lindsey, Gaska, Conley, Shapiro, Orlowski, Golden, Kaur, Sigh, Thelen, Laurenz, Davidson and Ciampitti2019) reported a reduction in seed oil and increases in soybean protein concentration in higher latitudes for soybean sowed in the USA. In the same way, Rotundo et al. (Reference Rotundo, Miller Garvin and Naeve2016) documented higher protein concentrations in lower latitudes in the northern hemisphere. Thakur and Hurburgh (Reference Thakur and Hurburgh2007) compared the quality of samples from different locations (in the USA and Argentina) and documented that Brazilian soybean had the highest seed oil and protein concentration. The reason for this higher protein concentration may be related to genetic breeding (flowering-time control and latitudinal adaptation, see Lu et al., Reference Lu, Zhao, Hu, Liu, Nan, Li, Fang, Cao, Kong, Su, Zhang, Li, Wang, Yuan, Cober, Weller, Liu, Hou, Tian and Kong2017) inherent to cultivars recommended for low-latitude regions. Yet, this has not enough been elucidated by science, more studies are required to advance our understanding of interactions between soybean protein and latitude.
Although the purpose of this study was not to test N-fertilization on soybean to make recommendations, it is important to clarify that N-fertilization applications of that magnitude can generate substantial volatilization losses that can exceed 40% (Cantarella et al., Reference Cantarella, Otto, Soares and de Brito Silva2018). These results alerted us to a considerable chance of increasing environmental losses under subtropical and tropical conditions by high rates of the volatilization of N-NH3 associated with urea application. Hence, this heavy fertilization practice (1000N) is not consistent with the sustainable intensification principles and is not cost-effective or environmentally appropriate – farmers should not consider the application of 1000 kg N/ha on soybean in tropical environments. Thus, improvements in N-management on soybean might increase protein production in Brazil and contribute to the global plant-based protein production for a growing population. However, our findings indicated that improvements in N-management on soybean might increase protein production in Brazil and contribute to the global plant-based protein production for a growing population.
Conclusions
Eleven soybean experiments were conducted under tropical and subtropical environments in Brazil with rainfed or irrigated conditions combined with N-fertilization treatment (0 or 1000 kg N/ha). The N-fertilization under subtropical and tropical conditions resulted in significant (in some environments under rainfed) or no soybean seed yield effect (under irrigation) and inconsistent seed oil concentration responses. However, our findings showed that the ample N supply in the 1000N treatment consistently increased seed protein concentration in the tropical and subtropical environments evaluated, and its magnitude was larger than what was previously reported for temperate environments. Finally, in lower latitude soybean regions of Brazil, the seed protein and oil concentration were higher than in higher latitudes.
This paper expands upon the work of the first author's PhD thesis.
Acknowledgements
We thank Daniel Medeiros from BASF Brazil for fungicides, insecticides, and herbicides donations for all experimental areas and Leonardo Gorgen from Yara Brazil for fertilizer donations for all experimental areas.
Author contributions
E. H. Figueiredo Moura da Silva conceived and designed the study, conducted data gathering in Piracicaba-SP, performed statistical analyses, wrote and edited the article. N. Cafaro La Menza wrote and revised the article. G. G. Munareto conducted data gathering in Cruz Alta-RS and Tupanciretã-RS. A. J. Zanon revised the article. K. Santos Carvalho conducted data gathering in Sorriso-MT. F. R. Marin conceived and designed the study, and revised the article.
Financial support
This research was supported by the Research Foundation of the State of São Paulo (FAPESP 2015/25702-3; 2017/23468-9; 2019/18303-6, 2017/20925-0, 2017/50445-0, 2021/00720-0, 2022/02396-8), Brazilian Research Council (CNPq, 301424/2015-2, 300916/2018-3, 401662/2016-0 and 425174/2018-2) and Coordination for the Improvement of Higher Education Personnel (CAPES) – Ministry of Education of Brazil.
Conflict of interest
None.
Ethical standards
Not applicable.








