



Introduction
Seed dispersal is a critical process for the spread and survival of plant populations (Janzen Reference Janzen1970; Nathan and Muller-Landau Reference Nathan and Muller-Landau2000; Nathan et al. Reference Nathan, Schurr, Siegel, Steinitz, Trakhtenbrot and Tsoar2008; Thomson et al. Reference Thomson, Letten, Tamme, Edwards and Moles2017). Fenner and Thompson (Reference Fenner and K2005) highlighted four advantages of seed dispersal in plants: (1) avoidance of specialist predators or pathogens of the parent plant, (2) mitigating the risk that seeds might face in highly heterogeneous or rapidly changing environments or landscapes, (3) minimizing competition with the parents or siblings, and (4) finding “safe sites” for successful germination and seedling establishment. Platt and Weis (Reference Platt and Weis1977) suggested that the likelihood of a plant species arriving in a new site depended on the number of dispersing propagules, the dispersal capacity of the propagules, and the distribution of the source plants. Species whose propagules include dispersal structures that facilitate long-distance dispersal by wind (e.g., comas, wings) are able to colonize novel environments even if most seeds often disperse close to the parent plant (Bullock and Clarke Reference Bullock and Clarke2000; McEvoy and Cox Reference McEvoy and Cox1987; Neubert and Caswell Reference Neubert and Caswell2000; Sheldon and Burrows Reference Sheldon and Burrows1973). For instance, Platt and Weis (Reference Platt and Weis1977) observed that in a guild of fugitive plant species colonizing badger (Taxidea taxus Schreber) mounds in an Iowa prairie, prairie dogbane (Apocynum sibiricum Jacq.) and common milkweed (Asclepias syriaca L.), which possess comose seeds, were most widespread on the site and had the greatest chance of reaching new mounds and preempting resources relative to species whose seeds had lower dispersal capabilities (e.g., hairy four o’clock, Mirabilis hirsuta (Pursh) McMill.; hoary vervain, Verbena stricta Vent.).
Management Implications
Black swallowwort [Vincetoxicum nigrum (L.) Moench] and pale swallowwort [Vincetoxicum rossicum (Kleopow) Barb.] are perennial, invasive vines of European origin in the dogbane (formerly milkweed) family. Both species are increasing in abundance in the northeastern United States and southeastern Canada. Prevention of spread is an important control strategy, especially because suppressing existing stands has proved difficult, labor-intensive, and sometimes ineffective. Information about the effects of wind speed, the height of seed release from the mother plant, and various seed-related characteristics can help inform prevention efforts. In our field and laboratory trials, we found that the release height of seeds and wind speed significantly affected dispersal distance in both Vincetoxicum species. Seeds of both species released from 2 m traveled greater distances than seeds released from 0.75 m, which fell within 20 m of the release point. The latter seeds would likely be responsible for consolidating already infested areas and expanding the invasion front. Vincetoxicum nigrum and V. rossicum seeds traveled up to 72 m and 79 m, respectively. These seeds are the most likely to colonize new areas and expand an infestation. To most effectively prevent spread to new areas and habitats, land managers should focus management efforts on plants growing farthest from the ground and where strong wind speeds are common, such as plants growing on the sides of gullies or twining up into trees. While these areas may be difficult to access, even one clipping of these plants before seed set would likely result in substantial reductions in the long-distance dispersal of seeds. Regular monitoring of these areas will be important so that any new populations are detected early enough to manage them effectively. Certainly, given that the vast majority of seeds in our study traveled no more than 20 m from the release point, management efforts should also focus on reducing seed production in established populations.
The distance traveled by seeds with special appendages such as a coma or plumes is in part affected by plant growth form, seed mass, the vertical height at which seeds are released, terminal velocity, and wind direction and speed at the time of seed release (Matlack Reference Matlack1987; Savage et al. Reference Savage, Borger and Renton2014; Tamme et al. Reference Tamme, Götzenberger, Zobel, Bullock, Hooftman, Kaasik and Pärtel2014; Thomson et al. Reference Thomson, Moles, Auld and Kingsford2011). The rate of propagule descent (i.e., terminal velocity) has often been related to the weight of the propagule and its size; this ratio has been referred to as plume or wing loading (Matlack Reference Matlack1987). Generally, the lower this ratio, the greater the likelihood that propagules will be transported greater distances at a given wind speed than propagules with relatively higher plume-loading ratios (Augspurger Reference Augspurger1986; Green Reference Green1980; Matlack Reference Matlack1987). The distance traveled is related to terminal velocity or settling rate, because propagules falling slowly are more likely to be exposed to horizontal winds and carried farther from source plants (Matlack Reference Matlack1987). Platt and Weis (Reference Platt and Weis1977) found that propagules of A. syriaca (terminal velocity=24.9 cm s−1) traveled an average of 13.8 m from source plants in an Iowa prairie at wind speeds of 10 to 15 km h−1, whereas propagules of A. sibiricum (terminal velocity=9.9 cm s−1) traveled an average of 25.7 m from source plants at the same wind speeds. Matlack (Reference Matlack1987) estimated that A. syriaca propagules (terminal velocity=27 cm s−1) could travel 18 m from source plants 2-m tall at wind speeds of 10 km h−1, whereas hemp dogbane (Apocynum cannabinum L.) propagules (terminal velocity=15 cm s−1) could travel 83.3 m at similar wind speeds. Sheldon and Burrows (Reference Sheldon and Burrows1973) suggested that the efficiency of propagule wind dispersal was determined to a greater extent by the fine details of plume or pappus geometry than by the ratio of the propagule weight to plume diameter or area. Regardless of the effect of plume geometry, plant species whose propagules have structures facilitating wind dispersal will likely be able to attain more distant sites and colonize new areas than species whose propagules lack such structures.
Wind-dispersed seeds released from taller plants are likely to travel farther than similar seeds released from shorter plants at the same wind speeds (Marchetto et al. Reference Marchetto, Jongejans, Shea and Isard2010b; Sheldon and Burrows Reference Sheldon and Burrows1973; Soons et al. Reference Soons, Heil, Nathan and Katul2004; Tackenberg et al. Reference Tackenberg, Poschlod and Bonn2003a; Thomson et al. Reference Thomson, Letten, Tamme, Edwards and Moles2017; Travis et al. Reference Travis, Smith and Ranwala2010). Herbaceous vines can overcome their lack of support structures and thus attain greater heights and growth by climbing up taller adjacent vegetation such as trees or other inanimate structures (Gianoli Reference Gianoli2015). This growth strategy not only allows herbaceous vines to escape competition but also allows their wind-dispersed propagules to travel much greater distances than would otherwise be the case if no tall nearby vegetation or other structures were present. Such a strategy may be especially important for vines that are able to colonize ecotone areas between forest and more open environments such old fields or pastures. Thus it is not surprising that 55 vines are listed as the world’s worst invasive species (Global Invasive Species Database 2013). Of these vines, 29% were introduced into North America from Asia, while only 2% are native to Europe (Sundarapandian et al. Reference Sundarapandian, Muthumperumal and Subashree2015). Of the top 10 worst invasive vines listed by Sundarapandian et al. (Reference Sundarapandian, Muthumperumal and Subashree2015) and occurring in the northeastern United States, only pale swallowwort [Vincetoxicum rossicum (Kleopow) Barb.] reproduces primarily by wind-dispersed seeds. Several of the other invasive species listed are woody perennial vines whose propagules are dispersed largely by birds and/or water [e.g., oriental bittersweet, Celastrus orbiculatus Thunb.; wintercreeper, Euonymus fortunei (Turcz.) Hand.-Mazz.]. All of these vines are aggressive growers and seed producers that can outcompete and displace native flora in invaded areas. Conceivably, all plant species, including herbaceous perennial vines such as V. rossicum that are highly dependent on sexual reproduction for range expansion, may benefit from having seeds adapted for long-distance wind dispersal, as this may increase the likelihood of reaching suitable and novel microenvironments (Cohen Reference Cohen1966; Salisbury Reference Salisbury1942).
Pale swallowwort [Vincetoxicum rossicum (Kleopow) Barb.=Cynanchum rossicum (Kleopow) Borhidi] and black swallowwort [Vincetoxicum nigrum (L.) Moench.=Cynanchum louiseae Kartesz & Gandhi] are herbaceous, perennial invasive vines in the Apocynaceae (dogbane and milkweed) family. Both species are self-compatible and either insect pollinated or self-pollinated (Lumer and Yost Reference Lumer and Yost1995; St Denis and Cappuccino Reference St Denis and Cappuccino2004). Introduced to the United States from Europe in the 1800s, these species have become increasingly problematic in the eastern United States and Canada (DiTommaso et al. Reference DiTommaso, Lawlor and Darbyshire2005b; Sheely and Raynal Reference Sheely and Raynal1996). Both Vincetoxicum species form dense stands and can dominate plant communities. In addition, V. rossicum has been reported as a problem in some perennial cropping systems such as Christmas tree farms, pastures, and no-till cropping systems (DiTommaso et al. Reference DiTommaso, Lawlor and Darbyshire2005b). Both species are capable of establishing in soils of varying pH and texture (Magidow et al. Reference Magidow, DiTommaso, Ketterings, Mohler and Milbrath2013; Sanderson et al. Reference Sanderson, Day and Antunes2015), and V. rossicum can readily germinate and establish across a range of temperatures (Sanderson and Antunes Reference Sanderson and Antunes2013).
Management of these Vincetoxicum species is difficult once populations have become established. Pulling or clipping of plants is often ineffective, and successful reduction of Vincetoxicum biomass usually requires repeated tissue removal or chemical control (Averill et al. Reference Averill, DiTommaso and Morris2008; DiTommaso et al. Reference DiTommaso, Milbrath, Bittner and Wesley2013; Milbrath et al. Reference Milbrath, DiTommaso, Biazzo and Morris2016). However, the use of herbicides may be undesirable, especially in natural areas, which are most often invaded. Cutting or mowing plants before seeds mature may reduce the number of seeds dispersed. However, not all invaded sites are easily accessible or suitable for cutting or mowing operations. Thus, significant research efforts have been made to find effective biological control candidates, with several European insects showing some promise (Milbrath and Biazzo Reference Milbrath and Biazzo2016; Weed et al. Reference Weed, Gassmann, Leroux and Casagrande2011).
Vincetoxicum nigrum and V. rossicum reproduce primarily via the production of high quantities of achenes (up to 32,000 m−2) that have comas to facilitate wind dispersal (Cappuccino et al. Reference Cappuccino, Mackay and Eisner2002; DiTommaso et al. Reference DiTommaso, Brainard and Webster2005a, Reference DiTommaso, Lawlor and Darbyshire2005b; Smith et al. Reference Smith, DiTommaso, Lehmann and Greipsson2006). If anecdotal reports of propagules attaching to the fur of white-tailed deer (Odocoileus virginianus Zimmerman) are confirmed, this may also aid dispersal. Plants also produce tillers from root crown buds, which allows for localized, short-distance expansion of populations (Averill et al. Reference Averill, DiTommaso, Mohler and Milbrath2011). Seeds of both Vincetoxicum species may be polyembryonic, although this feature is more prevalent in V. rossicum (Cappuccino et al. Reference Cappuccino, Mackay and Eisner2002; St Denis and Cappuccino Reference St Denis and Cappuccino2004; Smith et al. Reference Smith, DiTommaso, Lehmann and Greipsson2006) despite seeds of V. nigrum being generally heavier than those of V. rossicum (DiTommaso et al. Reference DiTommaso, Lawlor and Darbyshire2005b). While several studies have characterized Vincetoxicum seed dispersal, these have focused largely on the relationship between dispersal distance and seed size and the ability of these seeds to successfully establish (Cappuccino et al. Reference Cappuccino, Mackay and Eisner2002; Ladd and Cappuccino Reference Ladd and Cappuccino2005). Clearly, the establishment requirements of a plant species will define which sites are suitable and their availability across the landscape will determine the importance of dispersal ability (Janzen Reference Janzen1970; Platt and Weis Reference Platt and Weis1977). However, seed-dispersal distance is also influenced by height of seed release and horizontal wind speed (Dauer et al. Reference Dauer, Mortensen and Humston2006; Greene and Johnson Reference Greene and Johnson1989; Tamme et al. Reference Tamme, Götzenberger, Zobel, Bullock, Hooftman, Kaasik and Pärtel2014; Van Dorp et al. Reference Van Dorp, van den Hoek and Daleboudt1996). Given the wide range of habitats in which these two Vincetoxicum species occur (e.g., old fields, pastures, ecotones, forest understories), height of seed release and wind speed in addition to other seed traits (i.e., seed mass, settling rate) may be important in determining seed-dispersal distance and overall ability to spread to new environments.
The objective of this study was to determine the importance of seed release height, wind speed, seed mass, and settling rate (i.e., the rate of vertical fall in an undisturbed column of air) on the distance traveled by seeds of these two Vincetoxicum species. We hypothesized that increasing wind speed and greater release height will result in increased distance traveled by propagules and that greater plume loading (ratio of seed mass to coma diameter) will increase settling rate, which in turn will reduce the distance traveled. Seeds with slower settling rates are predicted to travel farthest because of increased time for horizontal wind to disperse them. Given that seed mass is generally greater for V. nigrum seeds relative to V. rossicum seeds and assuming that coma diameter is similar in these two congeners, we predicted that propagules of V. rossicum would, on average, travel greater distances than propagules of V. nigrum, suggesting that range expansion in V. rossicum may be of greater concern. Ultimately, the long-term objective of this research was to accurately determine dispersal distances for propagules of these two invasive vines. In turn, this would allow identification of habitats or regions that may be potentially reached and colonized by these species so that proactive management strategies could be implemented as early as possible.
Materials and Methods
Experimental Design
Vincetoxicum nigrum seeds were collected from Iona Island in Bear Mountain State Park, Rockland County, NY (41.30°N, 73.98°W) in August 2013 and 2015 and V. rossicum seeds from the Edwards Lake Cliffs Preserve in Ithaca, NY (42.52°N, 76.52°W) in September 2013 and 2015. Mature follicles (those open and dehiscing seeds or appearing partially dried and soon to open) were collected from a number of different plants, placed in paper bags, and allowed to dry at room temperature for several days. Undamaged seeds were then collected from the dried, open follicles. Seed-dispersal data were collected in two open environments near Ithaca, NY. Seeds of both species were released at both sites on multiple days during autumn 2013 and 2015 to maximize the range of wind speeds occurring during dispersal events (Table 1). Because wind speeds were directly measured, testing for differences between sites or days was not done.
Table 1 Number of seeds released by species per date at each study site.

Seed-Dispersal Distance
A seed-release apparatus was constructed by mounting a wooden clothespin to the top of a PVC pipe of length 0.75 or 2 m, which was then placed vertically over rebar driven into the ground so that the bottom of the pipe rested at ground level. A seed was placed individually in the clothespin, held by the smallest possible section of coma to avoid any damage. A string attached to the clothespin allowed a researcher to crouch down and pull to release the seed from a release height of 0.75 m or 2 m while minimizing wind disturbance near the seed. Seeds were followed until they came to rest, the location marked, and the distance traveled from the release point measured. Wind speed data were collected at the time of each release with a handheld anemometer (Kestrel 2000 Wind Meter, Nielsen-Kellerman, Boothwyn, PA 19061) at approximately 1-m height. A wind-profile power law relating wind speed at one height to wind speed at a second height suggests that wind speed increases less than 1 m s−1 between 0.75 and 2 m (Justus and Mikhail Reference Justus and Mikhail1976). Those seeds with intact comas following the dispersal study were collected for use in a subsequent laboratory study to assess their settling velocity (see “Settling Rate” section). In total, 151 V. nigrum seeds were released (Table 1), 65 at 0.75-m release height and 86 at 2 m. A total of 146 V. rossicum seeds were released (Table 1), 61 at 0.75 m and 85 at 2 m.
Settling Rate
Settling velocities of all recovered seeds with intact comas were measured following the protocol described by Andersen (Reference Andersen1992). Individual seed–coma units were dropped down a 1.22-m clear plastic tube with 8-cm diameter. Static Guard (Static Guard, B & G Foods, Parsippany, NJ 07054) spray was applied to the inside of the tube to prevent the seeds from clinging to the tube due to static electricity. Air movement inside the tube was presumed to be minimal. Three replicate measurements of falling time were obtained for each seed. Falling time in seconds was converted to mean settling velocities in meters per second. Seeds were then weighed, excluding the coma. The mass of seeds used in the dispersal study that had lost their comas was also determined. Settling-rate data were collected for 89 V. nigrum seeds (those retaining intact comas following the dispersal study), 43 from the 0.75-m treatment and 46 from the 2-m treatment. Settling-rate data were collected for 100 V. rossicum seeds, 35 from the 0.75-m treatment and 65 from the 2-m treatment. Although, at the time of the seed-dispersal distance and settling-rate studies, we did not measure the diameter (cm) of the coma for each seed, we were able to measure the diameter (cm) of fully expanded comas for 30 randomly selected V. nigrum seeds and 30 V. rossicum seeds from these same populations that had been carefully stored in open, paper bags at room temperature for 24 mo. Because most of the comas had detached from seeds during this storage period, we were unable to measure coma diameter and the weight of its corresponding seed. Thus, we calculated the plume-loading ratio for each of the species by comparing the average seed weight of the 30 seeds to the average diameter of the 30 comas. Marchetto et al. (Reference Marchetto, Jongejans, Jennis, Haner, Sullivan, Kelly and Shea2010a) reported that storing musk thistle (Carduus nutans L.) capitula in open paper bags for as long as 5 yr did not affect several seed traits (e.g., pappus damage, seed mass) or parameters measured, including terminal velocity. Thus, despite the low number of samples used and possible influence of this extended storage on coma structure, these data allowed us to determine whether there were any broad differences in the plume loading between the two species that could help explain dispersal-distance differences between them.
Statistical Analyses
All analyses were completed using R software (R Core Team 2015). Graphs were plotted with ‘ggplot2’ (Wickham Reference Wickham2009). Kernel density curves of seed dispersal were plotted for both species by release height. Various distributions were fit to the observed dispersal distances using the packages ‘SuppDists’ (Wheeler Reference Wheeler2013), ‘MASS’ (Venables and Ripley Reference Venables and Ripley2013), and ‘fitdistrplus’ (Delignette-Muller and Dutang Reference Delignette-Muller and Dutang2014). A lognormal distribution was found to have the best fit after the Akaike information and Bayesian information were compared (Supplementary Material S1). Data were log10-transformed, and a linear model was used to assess the effects of species, release height, wind speed, and all interaction terms on seed-dispersal distances. The linear model was subjected to an ANOVA with the ‘Car’ package (Fox and Weisberg Reference Fox and Weisberg2011).
Quantile regressions were performed for both species separately to investigate the effect of wind speed and height on seed dispersal of each quantile (also called tau) of distance with the ‘quantreg’ package (Koenker Reference Koenker2013). In ecology, quantile regression has been proposed and used as a way to determine more useful predictive relationships between variables in cases in which there is no relationship or only a weak relationship between the means of such variables (Cade and Noon Reference Cade and Noon2003). For example, mean seed dispersal, that is, the 0.50 quantile, is less relevant to the aim of this study than the 5% of seeds that dispersed the farthest, that is, the 0.95 quantile. Quantile regressions were implemented for tau from 0.05 to 0.95, by 0.05 increments, as distance=height+wind speed.
Differences in settling rates and seed mass by species were tested by t-tests. The relationships between seed mass and settling rates were depicted with smooth lines and a 95% confidence interval with ‘ggplot2’ for three categories of seed-dispersal distances: short, the 25% lowest values (1st quartile); long, 25% highest values (4th quartile); and mid, intermediate distances from 2nd to 3rd quartile.
Interactions between wind speed, representing horizontal movement, and settling rates, representing vertical wind-free movement, were investigated for their effects on seed-dispersal distance for both species with contour plots based on the linear model log10(distance)=settling rate+wind speed+settling rate×wind speed. Contour plots were generated with the contour function of the ‘rsm’ package (Lenth Reference Lenth2009).
Results and Discussion
Seed-Dispersal Distance
Main Explanatory Factors
Species, release height, wind speed, and the interaction between release height and wind speed had a significant effect on dispersal distances (Table 2). Overall, V. nigrum seeds landed closer to the release point than V. rossicum seeds, and seeds of both species released from 0.75 m were more likely to land closer to the release point (V. nigrum: 4.4±3.3 m, V. rossicum: 4.7±3.5 m; reported as mean±standard deviation) than seeds released from a 2-m height (V. nigrum: 12.6±9.6 m, V. rossicum: 17.1±12.1 m; Figure 1). The probability of a seed released from 0.75 m falling within 20 m of the mother plant was 100%, whereas 80.7% of seeds released from 2 m would do the same (Figure 1). Long-distance dispersal events (>20 m) only occurred for seeds released from 2 m (Figure 1)—the maximum distance measured in this study was 72.1 m for V. nigrum and 79.6 m for V. rossicum. These findings are comparable to results from an experiment using A. syriaca, the closest relative for which data are available, in which seeds traveled farther when released from higher points (Morse and Schmitt Reference Morse and Schmitt1985), and with the general trend that seeds produced by tall plants disperse greater distances than seeds from short plants (Thomson et al. Reference Thomson, Letten, Tamme, Edwards and Moles2017). Matlack (Reference Matlack1987) estimated that propagules of A. syriaca and A. cannabinum released from a height of 2 m could travel 18.1 and 83. 3 m, respectively when subjected to winds of 10 km h−1. Platt and Weis (Reference Platt and Weis1977) measured the distance traveled by 200 propagules each of A. syriaca and A. sibiricum in a Iowa prairie at wind speeds of 10 to 15 km h−1and reported mean distances traveled of 13.8 and 25.7 m, respectively. These workers did not specify the height from which propagules were released, except to state that they released seeds at the height they occurred on parent plants. However, A. syriaca plants can attain heights of 0.6 to 1.5 m (Bhowmik and Bandeen Reference Bhowmik and Bandeen1976) and A. sibiricum plants can grow as tall as 1.2 m, but are often 0.6- to 0.9-m tall (Minnesota Wildflowers 2018).

Figure 1 Kernel density curves of the seed dispersal of Vincetoxicum nigrum and Vincetoxicum rossicum according to release height (0.75 m or 2 m). The area under the curve of a density function represents the probability of finding a seed dropped from a particular height at a particular distance from the mother plant.
Table 2 Effects of species (Vincetoxicum nigrum and Vincetoxicum rossicum), seed release height (0.75 m and 2 m), and wind speed (km h−1) on the seed dispersal (distance in m, log10-transformed) assessed by linear model.

While all the 297 seeds tracked in this study were followed until they came to rest, several seeds released from the 2-m height in preliminary trials were lost. These seeds either traveled faster than researchers could track them or were carried high enough that they were lost from sight, usually at distances greater than 100 m from the release point. Cappuccino et al. (Reference Cappuccino, Mackay and Eisner2002) also reported losing sight of a V. rossicum seed released from a height of 1.5 m. Thus, for both species, rare events of very long-distance dispersal can occur under windy conditions, especially when seeds are released from heights of 1.5 m or greater.
Wind Speed versus Release Height for Short and Long Dispersal Distance
Because of the highly significant interaction between wind speed and release height (Table 2), quantile regressions were used to explore these two factors. Quantile coefficients estimated by quantile regression (Figure 2) show that release height was only important in explaining dispersal distances for both V. nigrum and V. rossicum for quantiles above 0.80; that is, release height was highly influential in promoting long-distance dispersal of Vincetoxicum seeds. Wind speed had little effect on dispersal of V. nigrum seeds until the 0.95 quantile (the longest distances recorded), where wind speed became highly predictive of dispersal distances. In contrast, wind speed was useful for predicting V. rossicum dispersal distances for the lower 85% of distances measured in our data set but was not predictive for the longest distances (Figure 2). Thus, for long-distance dispersal, both seed release height and wind speed were very important factors for V. nigrum, whereas only height appeared to matter for V. rossicum. A possible explanation for these findings is that for the generally heavier seeds of V. nigrum (see “Settling Rates and Relationship with Seed Traits” section), both higher seed release points and stronger winds were required for long-distance dispersal, whereas for the lighter V. rossicum seeds, long-distance dispersal could be achieved as long as they were at sufficient height to perhaps be caught up in updrafts and be transported in the atmospheric boundary layer (Savage et al. Reference Savage, Borger and Renton2014; Shields et al. Reference Shields, Dauer, Vangessel and Neumann2006; Skarpaas et al. Reference Skarpaas, Auhl and Shea2006). That height may be especially important for long-distance dispersal of V. rossicum seeds is also consistent with our finding that mean plume-loading ratio (i.e., seed weight:coma diameter) was greater for V. nigrum (2.6) than V. rossicum (1.8), thus possibly limiting long-distance dispersal of V. nigrum in the absence of strong winds even at relatively high propagule release heights (e.g., 2 m).
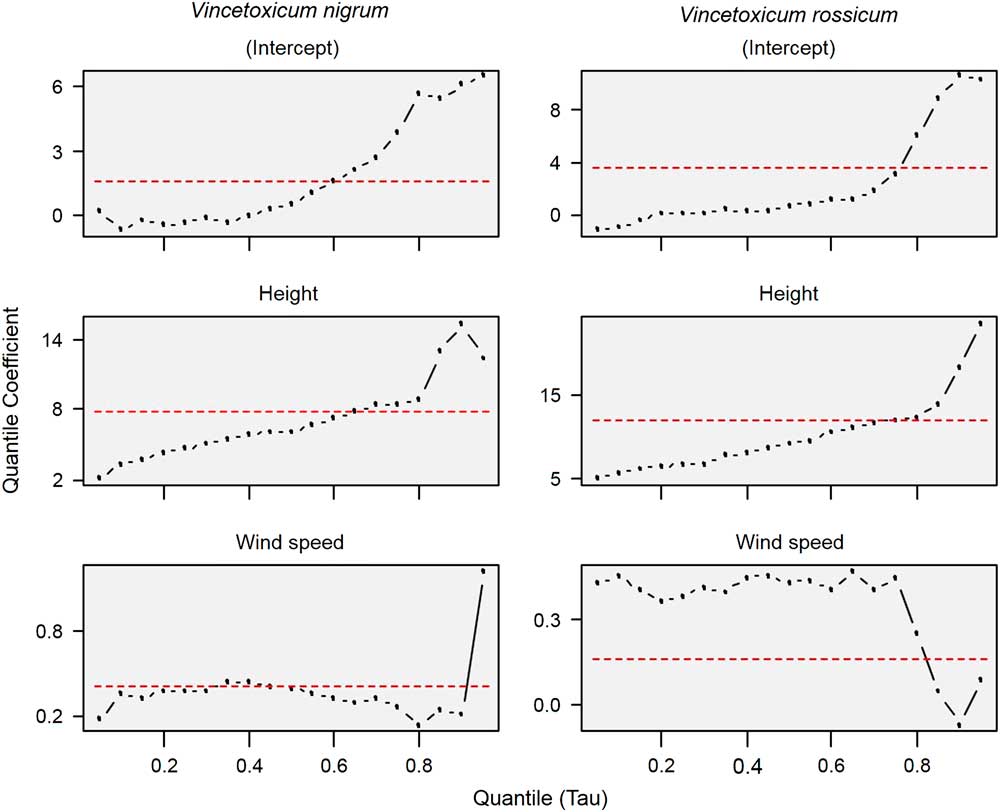
Figure 2 Quantile coefficients estimated by quantile regression (distance ~ height+wind speed) for each quantile (tau) of distance varying from 0.05 to 0.95. Red lines are the least-squares estimates. Dots are the value of the quantile coefficients (y-axis) for the particular tau value (x-axis).
The effect of wind speed on seed-dispersal distances for each species and release height was further examined with quantile regressions by selected quantiles (Figure 3). At 0.75-m height, the effect of wind speed on seed dispersal was primarily positive for both species, but differed among quantiles. Dispersal distance of V. nigrum seeds increased weakly with increasing wind speed but sharply increased at the 0.95 quantile, promoting rare long-distance dispersal (Figure 3A). For V. rossicum, dispersal distance increased significantly with increasing wind speed beginning at short distances, but this relationship was not evident for quantiles greater than 0.90 (Figure 3B). Neither V. rossicum nor V. nigrum dispersal distance was much affected by wind speed at the 2-m height, except for a weak effect on V. nigrum at the 0.95 quantile (Figure 3C and D). Wind speed was therefore more influential for seed dispersal when seeds were released at lower heights (0.75 m).
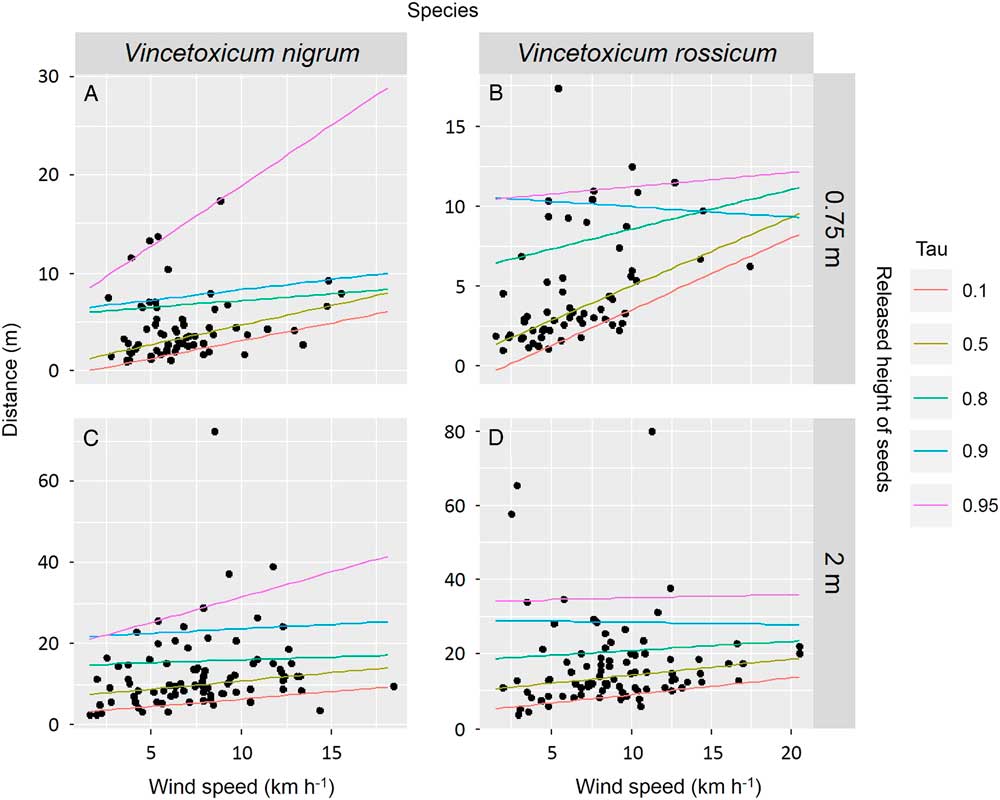
Figure 3 Quantile regressions for some selected quantiles (taus) predicting seed dispersal according to wind speed, by species (A and C, Vincetoxicum nigrum; B and D, Vincetoxicum rossicum) and release height. Dots are measured data. Lines are regression lines for each quantile (tau).
The limitations of measuring wind in the field must be noted. Wind-speed measurements reflected the conditions at the moment of propagule release, but it was not possible to measure all forces exerted on any propagule for the duration of its dispersal. While higher wind speeds were generally found to positively affect dispersal distance, making it more likely that a propagule would travel horizontally while it fell to the ground, wind is rarely a direct and steady horizontal force. Modeling studies have found that updrafts are an important factor affecting dispersal distances in dandelion (Taraxacum officinale G. H. Weber ex Wiggers) (Tackenberg et al. Reference Tackenberg, Poschlod and Kahmen2003b) and that turbulence is an important factor in long-distance dispersal (Nathan et al. Reference Nathan, Katul, Horn, Thomas, Oren, Avissar, Pacala and Levin2002; Savage et al. Reference Savage, Borger and Renton2014; Soons et al. Reference Soons, Heil, Nathan and Katul2004). It is probable that updraft and turbulence sometimes influence Vincetoxicum seed dispersal, but this is difficult to quantify in the field, and frequency of updraft events could vary according to the surrounding landscape. The few Vincetoxicum seeds previously mentioned that were lost in preliminary studies did appear to be caught by updrafts (CAS, personal observation).
Settling Rates and Relationship with Seed Traits
Mean settling rate was 0.59±0.23 m s−1 for V. nigrum and 0.43±0.11 m s−1 for V. rossicum, a significant difference by species (Figure 4A). Vincetoxicum nigrum showed a higher range of settling rates (from 0.23 to 1.43 m s−1) compared with V. rossicum (from 0.19 to 0.74 m s−1; Figure 4A). The faster settling rates of V. nigrum likely resulted in less time for potential effects of horizontal wind to occur, producing the shorter dispersal distances discussed earlier. Seed mass was also significantly higher for V. nigrum (0.013±0.008 g seed−1) compared with V. rossicum (0.008±0.004 g seed−1; Figure 4B) as was plume-loading ratio, as discussed earlier. Linear regression indicated that settling rate (SR) increased with increasing seed mass (SM) for V. rossicum (SR=42.3 SM+0.13; P<0.001; R2=0.44) and V. nigrum (SR=35.1 SM+0.14; P<0.001; R2=0.20). This relationship is what would logically be expected, as heavier seeds would likely fall faster, assuming comas of similar diameter (i.e., increasing plume-loading ratios). However, a further division of seeds into long- (25% highest values), moderate- (middle 50% values), and short-dispersing (25% lowest values) seeds did not always show the same seed mass by settling-rate relationships (Figure 5). In V. rossicum, as expected, settling rate increased in all three seed-dispersal divisions as seed mass increased. Cappuccino et al. (Reference Cappuccino, Mackay and Eisner2002) also predicted that heavier V. rossicum seeds would travel shorter distances from the source plants. In our study, the settling rate did not vary with seed mass for V. nigrum seeds that dispersed short distances, whereas for seeds that dispersed long and middle distances, settling rates increased with increasing seed mass and then reached a plateau. Because both small and high-mass seeds fell near the release point, the plume area of the coma may be more variable in short-dispersing V. nigrum seeds (Greene and Johnson Reference Greene and Johnson1990). Although we did not correlate coma diameter of V. nigrum seeds to the distance traveled, coma diameter in this species was much more variable (4.0 to 5.8 cm; mean=5.2 cm) in the 30 samples we measured compared with coma diameter for V. rossicum (4.3 to 4.7 cm; mean=4.5 cm).
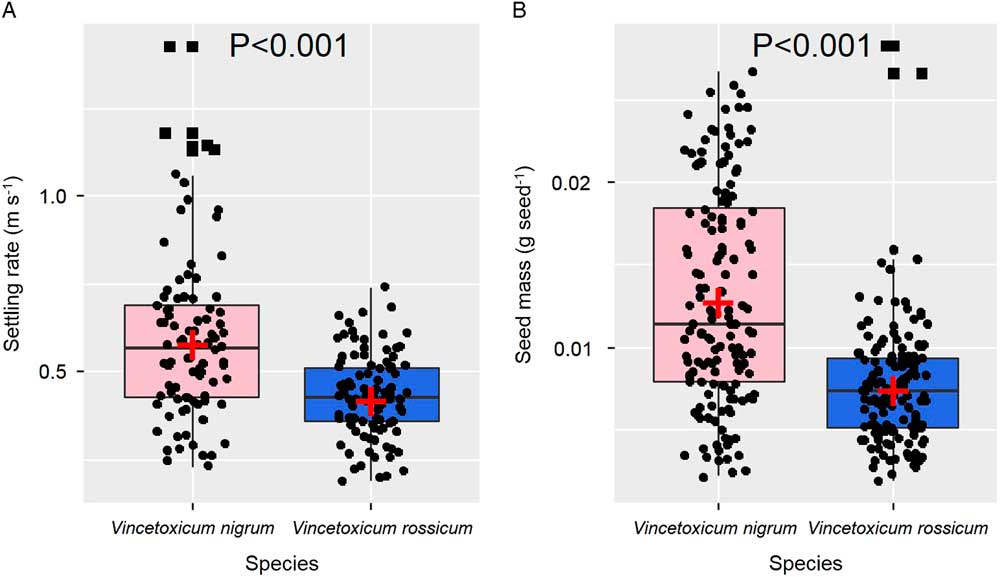
Figure 4 Settling rates (A) and seed mass (B) by species. Red crosses indicate averages. Black dots and squares are measured values; squares are values greater than 1.5× the box height (outliers). t-Tests for significant differences between species (P-value).

Figure 5 Relationship between seed mass and settling rate for seeds found at the shortest (Short: 25% lowest values, 1st quartile), longest (Long: 25% highest values, 4th quartile), and intermediate distances (Mid: from 1st to 4th quartile).
Regarding V. rossicum, two previous studies found that increasing seed mass resulted in shorter dispersal distances and greater germination and seedling survival (Cappuccino et al. Reference Cappuccino, Mackay and Eisner2002; Ladd and Cappuccino Reference Ladd and Cappuccino2005), although the authors noted that the relationship between mass and distance was weak. Ladd and Cappuccino (Reference Ladd and Cappuccino2005) suggested that if smaller seeds do indeed have decreased germinability and survival, coupled with a higher probability of long-distance dispersal, then long-distance dispersal in Vincetoxicum may not contribute as much to population spread as expected. However, in a 2-yr field study at two locations in central New York State, Averill et al. (Reference Averill, DiTommaso, Mohler and Milbrath2010) found that in the better-drained site, seeds of V. rossicum were able to germinate and seedlings to successfully establish in plots subjected to a range of disturbances. Disturbance regimes ranged from intact resident vegetation (controls) to highly disturbed plots in which the resident vegetation was killed using an herbicide followed by a tillage operation. Although establishment rates in their study were relatively low (15%±1%), these data nonetheless suggest that seeds of V. rossicum that may be dispersed long distances are capable of germinating and establishing seedlings in a wide range of heterogeneous microenvironments.
Trade-Off between Horizontal and Vertical Dispersal Speed
When seed-dispersal distances were estimated using a linear model including settling rate and wind speed, V. nigrum seeds were predicted to travel farthest when settling rate was low or wind speeds were high (Figure 6). In combination, lower vertical seed speeds (settling rate) and higher horizontal wind speed led to high seed dispersal. However, seeds with settling rates less than 0.3 m s−1 were predicted to disperse more than 10 m regardless of wind speed, and at wind speeds greater than 13 km h−1, predicted seed dispersal was 10 m or more regardless of the settling rate. Vincetoxicum rossicum showed similar trends, except when wind speeds were between 10 and 15 km h−1 and settling rates were lower than 0.3 m s−1 (Figure 6). Also, when wind speed exceeded 15 km h−1, seeds of any settling rate were predicted to disperse very far, particularly those with high settling rates (Figure 6). The response of V. nigrum is what would be expected: the slower a propagule falls, the more opportunity there is for wind to move the propagule horizontally before it comes to rest on the ground. The response of V. rossicum was also largely what was expected, except for the increased distance traveled at high wind speeds and high settling rate. It may be that these propagules have smaller coma diameters, which we did not measure, resulting in a higher plume-loading ratio and hence settling rate (Greene and Johnson Reference Greene and Johnson1990). Further experiments could examine the relationship of coma size or plume area and distance traveled, coupled with the other variables included here.

Figure 6 Contour plot representing seed dispersal (numbered lines are distances, back-transformed from log-10), estimated according to the horizontal wind speed and the vertical falling speed without wind (settling rate) by linear model as: log10(distance) ~ settling rate+wind speed+settling rate×wind speed.
Long-distance dispersal events of any plant are of great interest when considering possible establishment of new populations and spread of the species (Nathan Reference Nathan2006). Seeds landing very near the mother plant may bolster and slowly expand an existing population, but long-distance dispersal events have the potential to result in comparatively rapid colonization of a new area by a plant species. The potential for long-distance dispersal of propagules also reduces the likelihood that established seedlings in the new environment are negatively affected by specialist diseases or predators of the parent plants (Fenner and Thompson Reference Fenner and K2005; Janzen Reference Janzen1970). Because both Vincetoxicum species are self-compatible and either insect pollinated or self-pollinated (Lumer and Yost Reference Lumer and Yost1995; St Denis and Cappuccino Reference St Denis and Cappuccino2004), there is reduced risk that plants establishing far from source populations would be isolated from nonvectored pollen sources. This understanding of spread, coupled with the information that prevention is usually the most time- and cost-effective management strategy, suggests that preventing long-distance dispersal events is important in slowing population spread of invasive plants. Because both V. nigrum and V. rossicum are vines with stems reaching 2-m lengths or more (DiTommaso et al. Reference DiTommaso, Lawlor and Darbyshire2005b), it is conceivable that large numbers of seeds abscise from follicles at heights of 1 to 2 m from vines climbing up trees. In upstate New York, V. rossicum can often be found growing in gullies or on outcrops, where even greater release heights could be obtained. If the greater height of these seeds means a higher likelihood of increased dispersal distance, land managers might focus their efforts on plants growing in high, exposed areas or climbing up into trees, other vegetation, or structures. Because high wind speeds also have a major effect on longer-distance dispersal of V. nigrum, managers could target sites that experience the highest wind speeds or gusts for management efforts. While long-distance dispersal events may not contribute as much to population growth of Vincetoxicum spp. due to poor germination and survival of the seeds (Ladd and Cappuccino Reference Ladd and Cappuccino2005), the ability of a few seedlings to successfully establish in a new area following long-distance dispersal is likely to provide an opportunity for these species to substantially expand their range (Averill et al. Reference Averill, DiTommaso, Mohler and Milbrath2010). Where prevention or reduction of spread is a priority, land managers should focus their management efforts on plants growing farthest from the ground and where higher wind is expected (Marchetto et al. Reference Marchetto, Jongejans, Shea and Isard2010b). The findings reported in this study also provide important information for developing species-specific dispersal-kernel models that will aid in better targeting management efforts across the landscape and help prioritize the optimal traits of potential biological control agents to be used for managing these highly aggressive perennial vines (e.g., Shea Reference Shea2004; Skarpaas and Shea Reference Skarpaas and Shea2007).
Acknowledgments
The authors would like to thank Cornell University’s Musgrave Research Farm and Farm Services for providing field sites, Jerry Shen and Christine Geeng for invaluable service helping to track seeds in the field, and R. J. Richtmyer and Scott Morris for technical support. Funding support for CAS from the Cornell University Graduate School and the Section of Soil and Crop Sciences, School of Integrative Plant Science, is gratefully acknowledged. This work was also financially supported by French ANR COSAC project (ANR-14-CE18-0007) and the Burgundy Region in France (FABER project Couv’Herbi). The authors thank Fabrice Dessaint and Benoit Ricci (data scientist and landscape ecologist, INRA, UMR1347 Agroécologie, Dijon, France) for valuable discussions on data analysis. The comments and suggestions provided by three anonymous reviewers and the Associate Editor to improve an earlier version of this article are gratefully acknowledged. No conflicts of interest have been declared.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2018.8










