



Introduction
The Australian tree Melaleuca quinquenervia (Cav.) S.T. Blake (Myrtaceae) has been the target of a multidecade management effort that utilized a combination of chemical, biological, physical, and cultural tactics (LaRoche Reference Laroche1998). This integrated approach was guided by the Melaleuca Management Plan developed by the South Florida Water Management District, the Florida Exotic Pest Plant Council, USDA-ARS, and others in 1990 and revised in 1994 and 1999 (Florida Exotic Pest Plant Council 1999). During this period, M. quinquenervia was transforming large areas of greater Everglades landscape from a mosaic of sawgrass [Cladium mariscus ssp. jamaicense (Crantz) Kük] prairies, pine (Pinus sp.) flatwoods, and wet prairies into dense monotypic forests of M. quinquenervia. This led to disruptions of native communities from changing fire regimes, alterations to nutrient cycles, modifications of hydrology, and suppression of native plant species (Center et al. Reference Center, Purcell, Pratt, Rayamajhi, Tipping, Wright and Dray2012; Gordon Reference Gordon1998; Martin et al. Reference Martin, Tipping and Sickman2009; Rayamajhi et al. Reference Rayamajhi, Pratt, Center, Tipping and Van2009).
One of the original challenges in managing this species was its large seedbank, held in permanent serotinous capsules within the canopy, with the seed released synchronously in response to perturbations such as treatments by herbicides or wildfires (Center et al. Reference Center, Purcell, Pratt, Rayamajhi, Tipping, Wright and Dray2012). Following the desiccation of seed capsules, massive amounts of seeds were deposited near the tree that eventually recruited dense populations of seedlings (Woodall Reference Woodall1982). This species is intermediately serotinous, whereby some seeds are continually released to take advantage of favorable, yet unpredictable, germination microsites (Lamont et al. Reference Lamont, Le Maitre, Cowling and Enright1991). A few seed capsules (about 12% per year) desiccate and open continuously, producing a lighter but constant seed rain (Tipping et al. Reference Tipping, Martin, Pierce, Center, Pratt and Rayamajhi2012; Woodall Reference Woodall1982). Seeds are very small, which aids their penetration of leaf litter to form a short-lived soil seedbank with a half-life of about 1 yr (Van et al. Reference Van, Rayamajhi and Center2005). Although only about 9% of the seeds are viable, a single large tree located within a dense stand retains up to 9 million viable seeds within its canopy (Rayamajhi et al. Reference Rayamajhi, Van, Center, Goolsby, Pratt and Racelis2002). Recruited seedlings are highly susceptible to drought or flooding (Lockhart et al. Reference Lockhart, Austin and Aumen1999; Tipping et al. Reference Tipping, Martin, Pierce, Center, Pratt and Rayamajhi2012).
Management Implications
The invasive capacity of Melaleuca quinquenervia may be reduced as evidenced by a greater than 95% reduction in the size of recruitment cohorts following wildfires in 1998 and 2007 in the same area. In the past, before the implementation of classical biological control in 1997, mass recruitment events post-fire promoted this species to dominate landscapes. Today, seedlings, saplings, and large trees are now attacked continuously in Florida by three biological control agents. The density of the 1998 recruits was reduced by 96.3%, and none had produced seed capsules by 2005. In addition to reducing the aerial seedbanks of reproductive trees, the biological control agents reduced the rate of growth and height of the recruited seedlings following the second fire in 2007. After 3 yr, the natural mortality of new recruits (89.4%) increased slightly to 96.6% when attacked by the biological control agents. Recruits were 63% taller during the same period when protected from the agents using insecticides. There was no recruitment during the next 3 yr despite seed rain and periodically favorable hydrological conditions. Newly recruited seedlings face not only high levels of natural mortality, but also constant herbivory from biological control agents, the combination of which will reduce their numbers, slow their growth, and inhibit the survivors from achieving the capacity to reproduce for many years. Based on the results of this study, land managers should target mature reproductive trees rather than cohorts of recruits to exert maximum suppression on M. quinquenervia populations.
The original role envisioned for the biological control portion of the Melaleuca Management Plan was to reduce this seed production and retard the growth of recruits. This goal was eventually accomplished with the establishment of three biological control agents that reduced seed production and plant growth rates, while increasing the susceptibility of saplings and smaller trees to wildfires and herbicides (Tipping et al. Reference Tipping, Martin, Pratt, Center and Rayamajhi2008, Reference Tipping, Martin and Gettys2017). The first agent, Oxyops vitiosa Moore (Coleoptera: Curculionidae), was released in 1997 and proved to be highly effective in areas that are not permanently inundated (Center et al. Reference Center, Van, Rayachhetry, Buckingham, Dray, Wineriter, Purcell and Pratt2000; Tipping et al. Reference Tipping, Martin, Nimmo, Pierce, Smart, White, Madeira and Center2009). This agent appears to promote a resource-regulation phenomenon that maintains a higher level of agent attack while reducing resources for reproductive tissues (Tipping et al. Reference Tipping, Martin, Pratt, Rayamajhi and Gettys2015). As a result of repeated defoliation, seed production is reduced by 80% to 100%, depending on conditions, and this loss of seed pressure appears to have influenced M. quinquenervia population dynamics to the point where the plant’s invasiveness is now reduced in certain areas (Tipping et al. Reference Tipping, Martin, Pratt, Center and Rayamajhi2008, Reference Tipping, Martin, Pierce, Center, Pratt and Rayamajhi2012).
The second agent, released in 2002, was Boreioglycaspis melaleucae (Hemiptera: Psyllidae), an agent unaffected by hydrology and with the demonstrated potential to suppress saplings and cause older trees to drop leaves (Morath et al. Reference Morath, Pratt, Silvers and Center2006; Purcell et al., Reference Purcell, Balciunas and Jones1997). This species established quickly and dispersed rapidly throughout the range of M. quinquenervia in Florida (Center et al. Reference Center, Pratt, Tipping, Rayamajhi, Van, Wineriter, Dray and Purcell2006). It appears to be temperature sensitive, with larger populations developing during the cooler dry season and smaller populations during the warmer rainy season (Chiarelli et al. Reference Chiarelli, Pratt, Silvers, Blackwood and Center2011).
The third agent was Lophidoplosis trifida Gagne (Diptera: Cecidomyiidae), a galling fly that was released in 2008 (Purcell et al. Reference Purcell, Wineriter and Brown2007). It also established readily and spread quickly, and its gall-forming activities can reduce sapling height by 10%, leaf biomass by 42%, woody biomass by 42.7%, and root biomass by 30.3% (Pratt et al. Reference Pratt, Rayamajhi, Tipping, Center, Wright and Purcell2013; Tipping et al. Reference Tipping, Martin and Gettys2016).
These combined cooperative efforts by multiple state and federal agencies using chemical, biological, physical, and cultural tactics have reduced M. quinquenervia infestations in both scale and density, leading to reductions in management costs, with concomitant increases in native plant diversity in formerly infested areas (Rayamajhi et al. Reference Rayamajhi, Pratt, Center, Tipping and Van2009). The impact of biological control on larger trees (>5-m height) growing in lower densities remains uncertain, given the difficulty in establishing true herbivory controls on trees of this size class, but flowering and seed production are often apparent in these trees. Some of these larger trees remain on the landscape in reduced densities and have led some land managers to target them out of concern they might serve as foci for reinvasion. These efforts can be expensive because of the need to deploy helicopters or ground crews to locate and kill isolated trees using herbicides.
The primary objective of this study was to evaluate the potential of these larger, solitary trees to serve as a seed source that might spur a reinvasion in areas largely cleared with chemical, biological, cultural, and mechanical methods. Secondarily, we measured the change in recruitment patterns following fires in 1998 and 2007 and the fate of recruited seedlings exposed to herbivory by the three biological control agents.
Materials and Methods
As part of an earlier experiment in 1998, 90 plots (0.5 m−2) were established within a 20-ha site in the Belle Meade section of Picayune State Forest (26.1023°N, 81.6422°W) to evaluate the impact of the recently released O. vitiosa on M. quinquenervia seedlings and saplings. The plant communities at this site, soil types, biogeochemistry, and hydrology were described by Martin et al. (Reference Martin, Tipping and Sickman2009). Researchers counted the number of recruits following the fire in December 1998 before the larger experiment was vandalized and subsequently abandoned.
In July 2005, six large solitary M. quinquenervia trees (mean height + SE: 12.4 + 1.2 m) growing in the same area with apparent capsule clusters visible in their canopies were selected for another experiment. Four plots (4 m2) were established within the seed shadow of each tree in cardinal directions, and all existing M. quinquenervia saplings (recruited after the 1998 fire) were counted, removed, and measured for above- and belowground biomass. A seed trap was installed adjacent to each quadrat to capture seed rain according to the design of Tipping et al. (Reference Tipping, Martin, Pierce, Center, Pratt and Rayamajhi2012). Once a month, the following data were collected from each plot: (1) number of seeds in seed traps, (2) the number and height of recruited seedlings in the quadrats, (3) water depth, and (4) soil moisture. Four soil samples (5-cm depth) were taken adjacent to the quadrats, weighed, and then dried to a constant weight to estimate soil moisture. During this experiment, O. vitiosa and B. melaleuceae were common at this site, while L. trifida spread into the plots near the end of the experiment.
In April 2007, the Great Balsa fire burned through all the plots and, for the first time since the 1998 fire, seedling recruitment occurred at the Belle Meade site, which led the researchers to modify the original experimental design to better evaluate the event (Florida Department of Forestry n.d.). A total of 975 post-fire recruited seedlings across all existing quadrats were numbered with a small metal stake and measured for height (Figure 1). The original quadrats around the reproductive trees were randomly assigned to one of two treatments so that half were sprayed with the insecticide acephate (OS-dimethyl acetylphosphoramidothiote) applied with a hand-pressurized backpack sprayer at a concentration of 0.367% ai (v/v) until wet (restricted herbivory) and the other half with water (unrestricted herbivory) every 4 to 6 wk. The data captured post-fire remained the same, with the addition of recording the number of surviving recruits and their heights periodically. If a seedling could not be located next to a numbered stake, it was considered dead, and the stake was removed. The study ended in May 2010, and all surviving recruits were harvested, separated into component parts (leaves, wood, roots, etc.), and dried to determine dry weight (DW) biomass.

Figure 1. Recruited Melaleuca quinquenervia seedling from the 2007 fire marked with a metal stake and tag.
Data Analysis
Data were tested for normality and homogeneity of variance and transformed using square-root transformation as needed before analysis. The mean mortality and height of the recruited cohort were calculated and used in a general linear model, with herbivory treatment as a fixed effect and sample date and sample tree as random effects (SAS Institute 2009). Direct comparisons of treatment effects on these final parameters were conducted with t-tests. The soil seedbank was estimated by assuming it was zero at the beginning of the experiment and cumulatively adding the seed rain, while subtracting recruited seedlings. Seed that fell into the plots was assumed to be nonviable after 2 yr and was subtracted from the estimated soil seedbank. Harvest biomass data from the 2007 cohort were subjected to one-way ANOVA, and the means between restricted and unrestricted herbivory treatments were separated using t-tests (SAS Institute 2009).
Results and Discussion
The mean number of recruits following the 1998 fire was 561.5 + 36.7 seedlings m−2 (TD Center, unpublished data) and by July 2005, the mean density (+SE) of the surviving saplings had declined 96.3% to 20.4 + 1.7 saplings m−2, none of which had any evidence of seed capsules. As mentioned earlier, these were then hand pulled to set up the second experimental plots around the individual trees. Following the establishment of the new plots, a total of two seedlings were recruited across all plots from July 2005 through March 2007 despite an average of 0.49 viable seeds m2 d−1 falling into the quadrats and periodically favorable hydrologic conditions (Figure 2). Seed rain appeared to increase beginning in May 2007, likely as a result of seed capsules desiccating after the fire and releasing seeds (Figure 2). Despite this, no seedlings were recruited for the next 5 mo until October 2007, when soil moisture and water levels increased to stimulate germination (Figure 2). After that pulse of recruitment between the end of September 2007 through mid-October 2007, no new recruitment occurred during the remaining 34 mo of the experiment, despite regular and progressively heavier seed rains (Figure 2) and an increasing estimated soil seedbank (Figure 3).
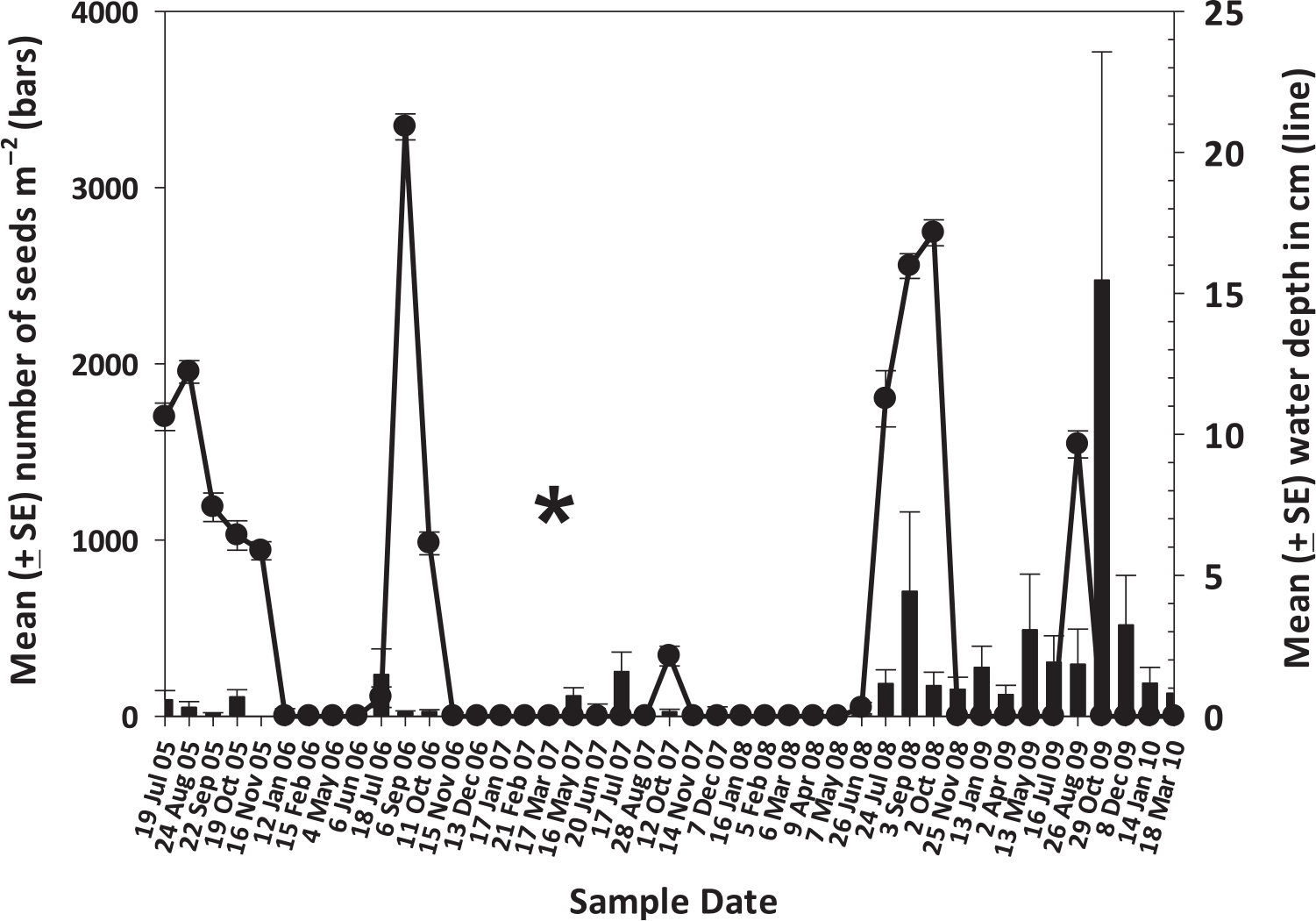
Figure 2. Mean seed rain (bars) from experimental Melaleuca quinquenervia trees with associated mean water depths (line) from July 2005 through March 2010 in the Belle Meade section of Picayune State Forest, Florida. The asterisk (*) indicates the approximate date of the 2007 Great Balsa fire.
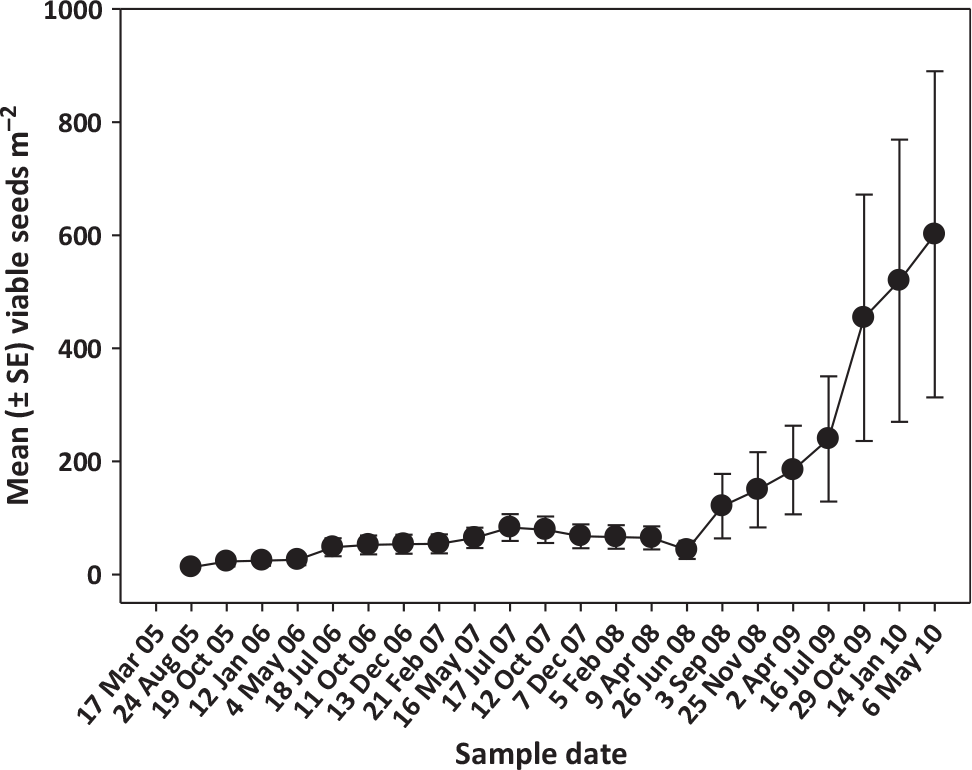
Figure 3. Mean estimated viable soil seedbank for Melaleuca quinquenvervia from August 2005 through May 2010.
The initial mean densities of the 2007 cohorts were equivalent in their assigned unrestricted and restricted herbivory treatments at 12.4 + 2.6 and 8.56 + 2.0 seedlings m−2, respectively (t = 1.13, df = 21, P = 0.26). Over the experimental duration of 1,083 d, the mortality trends of seedlings in both treatments were similar, and there was no difference in the final seedling mortality, with 96.6 + 1.3% of seedlings dying in the unrestricted herbivory treatment (water) and 89.4 + 4.9% dying in the restricted herbivory treatment (insecticide) (t = 1.47, df = 21, P = 0.15) (Figure 4). At the end of the experiment, the mean densities were also not different between treatments, with 0.27 + 0.07 saplings m−2 present in the unrestricted herbivory treatment and 0.97 + 0.44 saplings m−2 in the restricted herbivory treatment (t = 1.13, df = 21, P = 0.12). Those seedlings/saplings protected from herbivory grew taller than unprotected plants, with final mean heights of 101.3 + 2.2 cm under restricted herbivory and 37.4 + 6.9 cm under unrestricted herbivory (t = 8.15, df = 11, P = 0.0001) (Figure 5). Herbivory by the biological control agents reduced individual plant biomass by 90.1%, with a final mean DW biomass of 51.8 + 9.8 g under restricted herbivory versus 5.1 + 2.04 g under unrestricted herbivory (t = 5.04, df = 11, P = 0.0004). No flowers or seed capsules were produced by any of the recruits.

Figure 4. Mean cumulative mortality of Melaleuca quinquenervia seedlings recruited after the 2007 fire and subjected to unrestricted and restricted herbivory by biological control agents.

Figure 5. Mean periodic heights of Melaleuca quinquenervia seedlings recruited after the 2007 fire and subjected to unrestricted and restricted herbivory by biological control agents. *Treatment means within date are significantly different at P = 0.05.
The insecticide regime in this study was effective at controlling all three agents, including the more difficult to manage L. trifida, for which gall production was reduced by more than 87% (P = 0.02). Although some of the protected plants were stimulated to form galls, it is likely the flies within were killed by the insecticide treatment (Tipping et al. Reference Tipping, Martin and Gettys2016). The surviving recruits in the unrestricted herbivory treatment weighed 90.1% less (P = 0.0001) and were 63% shorter (P = 0.0001) than those protected with insecticides, but despite the apparent damage inflicted on the unprotected recruits by the biological control agents, they survived at the same level as protected recruits (P = 0.15).
Although the large M. quinquenervia trees in this study continued to disperse seeds between fires, the numbers were insufficient to recruit new populations without the intervention of a major perturbation like a fire. This is a common strategy in serotinous species, whereby wildfires influence both the quantity of available seeds plus improve the quality of the seedbed by burning off organic material to expose more mineral soils (Johnston and Elliott Reference Johnston and Elliot1998). Organic materials may promote high diurnal temperature fluctuations and lead to rapid drying of the soil surface (Johnstone and Chapin Reference Johnstone and Chapin2006). At seasonally inundated sites like the Belle Meade site, a dark-colored muck layer is left on the surface of the soil after the water recedes, and this layer dries out and forms a crust that oxidizes over time. The underlying mineral soils are typified by loamy, siliceous, hyperthermic Arenic Glassoqualfs (Pineda series), which can moderate temperatures while wicking moisture from subsurface layers to relieve the moisture limitation of seedlings like M. quinquenervia that are vulnerable to dry conditions (Johnstone and Chapin Reference Johnstone and Chapin2006; Martin et al. Reference Martin, Tipping, Reddy, Daroub and Roberts2010). Seedlings recruited atop this crust are likely to be at increased risk of drought stress as the crust dries, which may reduce their survival.
Before seed production was curtailed by O. vitiosa, M. quinquenervia trees maintained a large aerial seedbank that responded rapidly to perturbations like wildfires and exploited more favorable recruitment environments. With the 2007 fire, there was still a time lag of 5 mo until water levels were more favorable for seed germination before substantial recruitment occurred, and the recruitment window was relatively narrow, lasting only 2 mo. A characteristic feature of fire-prone environments is the tendency for recruitment to be concentrated in the first year following fire, at a time when resource availability is likely to be at its maximum (light, nutrients, soil moisture) and competition from established plants at its minimum (Lamont and Enright Reference Lamont and Enright2000). Previous studies at this site demonstrated the vulnerability of young seedlings to fluctuations in water stage, either from drying out or being drowned before they attained sufficient size (Tipping et al. Reference Tipping, Martin, Pierce, Center, Pratt and Rayamajhi2012). Thus, although biological windows still exist whereby reproductive M. quinquenervia can recruit seedlings following the combination of a perturbation and the right soil moisture conditions, the reservoir of seeds in the aerial seedbank appears to be much smaller, most likely as a result of years of herbivory by biological control agents. The estimated soil seedbank at this site appeared to be increasing almost a year after the 2007 fire, which may be explained by a pulse of nutrients that were scavenged by the large trees post-fire and eventually converted into reproductive biomass that resulted in an increase in inter-fire seed rain. Even though smaller seeds can penetrate inter-fire litter more easily and facilitate establishment, smaller seeds are often short-lived (Witkowski et al. Reference Witkowski, Lamont and Connell1991). In areas with more frequent fires (<20-yr intervals), serotiny has an advantage over soil storage, in which only a proportion of seeds will germinate, even if conditions are ideal (Lamont and Enright Reference Lamont and Enright2000). Where conditions are suitable for inter-fire establishment (such as the appearance of small gaps, exceptionally wet years), some spontaneous seed release may be advantageous (Lamont and Enright Reference Lamont and Enright2000). However, that did not appear to be the case at this site, where only two seedlings were recruited during the inter-fire portions of the experiment.
Factors that promote post-fire germination and seedling establishment may include the reduction or elimination of herbivore populations (Hanley and Fenner Reference Hanley and Fenner1997; Lamont et al. Reference Lamont, Le Maitre, Cowling and Enright1991). Although a number of studies, including this one, have identified the overall impact of herbivores on post-fire seedling populations using herbivore exclusion techniques, this kind of approach sheds little light on the actual mechanisms involved, including the early development and expression of seedling chemical or physical defenses (Hulme Reference Hulme1996; Karban and Thaler Reference Karban and Thaler1999). Regardless of the circumstances, it was clear in this case that the specialized herbivores were able to quickly locate and attack newly recruited seedlings.
Despite the post-fire pulse of recruitment in 2007, more than 96% of recruits were dead after about 3 yr, and the survivors will probably not flower for years to come, as evidenced by the recruitment cohort from 1998, for which none of the survivors had produced seed capsules after 7 yr. Plants protected from herbivory can reproduce within a year or two (Meskimen Reference Meskimen1962). Comparing differences in growth rates between appropriate experimental cohorts provides a rough estimate of 15 yr before any new recruits will achieve reproductive status because of the current levels of herbivory from biological control agents.
Seedling mortality from 1998 to 2005 matched that from 2007 to 2010 (93.6%), with only a slight increase in mortality between protected and unprotected seedlings. Another study at the Belle Meade site determined that 99.2% of the recruited seedlings died after 3 yr (Tipping et al. Reference Tipping, Martin, Pierce, Center, Pratt and Rayamajhi2012). Edwards and Crawley (Reference Edwards and Crawley1999) and others have argued that any seedling losses from herbivory contribute little to the plant community structure because of density-dependent mortality from intraspecific competition as a seedling cohort undergoes self-thinning. If this is true, the real impact of herbivory is not with seedling mortality but rather seed reduction and storage in reproductive M. quinquenervia, along with a delay between the emergence of a seedling and its development into a reproductive tree. The longer-term population-level implications of these differences are unknown but may suggest that the biological control agents will inhibit or delay the development of saplings to reproductive trees, as evidenced by the lack of seed capsule clusters on the 7-yr-old saplings/trees that were recruited after the 1998 fire. Another experiment conducted nearby during 2002 to 2007 using this same cohort found that trees began flowering about a year after the start of regular applications of insecticides, while no unsprayed trees ever flowered (Tipping et al. Reference Tipping, Martin, Nimmo, Pierce, Smart, White, Madeira and Center2009). There has been an overall decline in M. quinquenervia density at this site, as recruitment events have contracted in scale and scope, while the mortality of recruits remains high. This density reduction is concurrent with declines in seed production by individual trees caused by attack from biological control agents (Tipping et al. Reference Tipping, Martin, Pratt, Center and Rayamajhi2008). Land managers can consider this new reality when prioritizing management decisions; namely, high levels of seedling/sapling mortality without regular replacement and extended periods before M. quinquenervia recruits become reproductive because of herbivory by biological control agents.
Similar recruitment events have occurred nearby since this experiment, most recently in Big Cypress National Preserve, which is located east of the site of this experiment. In that case, areas adjacent to a few reproductive trees recruited seedlings following the 24,685 hectare Buzzard Fire (25.9762, 80.9408) that was sparked by lightning on April 10, 2018. As in this experiment, the seedlings were quickly attacked by biological control agents despite the low density and isolated nature of the mature M. quinquenervia (M Smith, personal communication). This confirms once again the ability of biological control agents to find isolated populations of seedlings and saplings and quickly build up their numbers.
Land managers at this preserve and Everglades National Park have used a variety of methods to control isolated M. quinquenervia, including spraying trees with herbicides delivered by drop nozzles slung under helicopters. Although this method requires specialized equipment and is expensive upfront (helicopter rental is about US$1,500 h−1), it reduces the overall costs of treatments in several ways, including reducing negative environmental impacts to the area by ground crews who must create or reopen paths to the infestations; it increases the likelihood of locating isolated trees; and it allows more areas to be treated more effectively in less time. For example, recent treatments in Big Cypress Preserve and Everglades National Park cost anywhere from US$18 to US$125 per tree, far less than the cost of a five-person ground crew at US$2,000 to US$3,000 h−1, assuming that ground crews could even access these areas and find the trees in the difficult terrain and dense vegetation. In the more heavily forested and roadless landscapes of Big Cypress National Preserve, a typical ground crew can normally locate and treat two sites a day (W Snyder, personal communication). In comparison, a helicopter with a drop nozzle can locate and spot-treat M. quinquenervia trees in an area as large as 5,666 ha in 2 d (H Cooley, personal communication; W Snyder, personal communication).
Although recently recruited M. quinquenervia populations may cause concern to land managers, prioritizing the elimination of reproductive trees may be the best use of scarce management dollars rather than spraying or burning recently recruited populations of seedlings/saplings, because the normally high levels of natural mortality from abiotic factors are no longer offset by seed rain, coupled with constant herbivory from biological control agents that retards the survivors’ growth and development toward reproductive capability. Any newly recruited sites can be mapped and left to the biological agents for at least a decade or longer before any trees that survive and produce seeds would need to be treated.
Acknowledgments
This research received no specific grant from any funding agency or the commercial or not-for-profit sectors. We thank Eileen Pokorny and Danyelle Fitzgerald for their assistance in sample collection and processing. We thank Ted Center for allowing us to use his unpublished data and Melissa Smith for discussions on her latest research. William Snyder of Big Cypress National Preserve and Hillary Cooley of Everglades National Park shared their insights and experience in managing M. quinquenervia. No conflicts of interest have been declared.





