


Liver and muscle glycogen are essential substrate sources during prolonged moderate-to-high-intensity exercise(Reference Coyle, Coggan and Hemmert1, Reference Bosch, Dennis and Noakes2). Hepatic glucose output increases in response to exercise(Reference Petersen, Price and Bergeron3, Reference Stevenson, Thelwall and Thomas4) to maintain circulatory glucose levels to meet the demands of increased substrate oxidation within skeletal muscle, especially late in exercise when muscle glycogen availability is reduced(Reference Bosch, Dennis and Noakes2). Both liver and muscle pre-exercise glycogen contents are directly associated with exercise performance(Reference Casey, Mann and Banister5, Reference Bergstrom, Hermansen and Hultman6). Therefore, the optimisation of short-term ( < 8 h) replenishment of both liver and muscle glycogen stores through nutritional intervention is potentially important for subsequent exercise performance, but remains unclear.
There is a positive relationship between the quantity of carbohydrate (CHO) consumed and muscle glycogen synthesis rates(Reference Jentjens and Jeukendrup7), reaching a ceiling of approximately 1·2 g/kg body mass (BM) per h, where muscle glycogen synthesis rates are approximately 9–10 mmol/kg wet weight (WW) per h(Reference van Loon, Saris and Kruijshoop8–Reference Jentjens, van Loon and Mann10). The CHO composition also influences the rates of muscle glycogen synthesis, with glucose (high glycaemic index) conferring benefits over CHO with a lowers glycaemic index, such as fructose(Reference Van Den Bergh, Houtman and Heerschap11). Glucose polymers, such as maltodextrin (MD), are also reported to promote approximately 25 % greater muscle glycogen synthesis compared with a mixture of glucose, maltose and lower-mass oligomers, due to increased gastric emptying and absorption(Reference Piehl Aulin, Soderlund and Hultman12).
The restoration of muscle glycogen post-exercise is driven by an initial rapid insulin-independent phase, lasting 30–60 min, followed by a longer insulin-dependent phase(Reference Price, Rothman and Taylor13). The postprandial insulin response can be enhanced by the addition of protein (PRO) to CHO, producing higher rates of muscle glycogen synthesis(Reference Zawadzki, Yaspelkis and Ivy14). PRO in combination with amino acids (AA), leucine and phenylalanine, of known insulinogenic potential(Reference van Loon, Saris and Verhagen15) may be of additional benefit to glycogen synthesis compared with PRO alone(Reference van Loon, Saris and Kruijshoop8). Post-exercise, the addition of PRO and AA to a moderate amount of CHO ( ≤ 0·8 g/kg BM per h) is reported to enhance muscle glycogen synthesis(Reference van Loon, Saris and Kruijshoop8), even when PRO displaces CHO from the supplement (i.e. isoenergetic(Reference Berardi, Price and Noreen16, Reference Ivy, Goforth and Damon17)). However, other reports have failed to reproduce these observations(Reference Tarnopolsky, Bosman and Macdonald18, Reference Betts, Williams and Boobis19). This may relate to the lower relative ingestion rate of PRO in these studies or the absence of supplementary AA, not enhancing the insulin response during recovery. The addition of PRO and PRO with AA to higher amounts of CHO than considered optimal ( ≥ 1·2 g CHO/kg BM per h) for muscle glycogen synthesis does not further enhance synthesis(Reference van Hall, Shirreffs and Calbet9, Reference Jentjens, van Loon and Mann10), even when the supplements are tested under isoenergetic conditions(Reference van Loon, Saris and Kruijshoop8, Reference Howarth, Moreau and Phillips20, Reference Rotman, Slotboom and Kreis21). The practical implication for muscle glycogen synthesis is that PRO added to CHO only appears to be of value when CHO supply is restricted. Thus, the type, dose and rate of supply of CHO become key determinants in glycogen synthesis in situations where prescription of PRO and AA has the potential to facilitate a time-dependent repletion. It remains to be determined if such a situation is true for the repletion of liver glycogen and whether adequate liver glycogen repletion is similarly attained by the use of CHO alone. Furthermore, the ecological relevance of enhanced muscle glycogen recovery in terms of endurance performance is yet to be fully established.
Despite the pivotal role of the liver during exercise, very little is known about the recovery of liver glycogen stores post-exercise, primarily due to the difficulty in taking biopsies from the liver for research purposes. This has been overcome by the use of non-invasive techniques, such as 13C magnetic resonance spectroscopy (13C-MRS), to evaluate liver glycogen. In vivo 13C-MRS is technically challenging due to the low natural abundance for this nucleus and its low MR sensitivity, though improved hardware and methodological approaches in recent years have provided valuable insights into liver glycogen dynamics(Reference Casey, Mann and Banister5, Reference Decombaz, Jentjens and Ith22). 13C-MRS has demonstrated that glucose and sucrose were equally effective at restoring liver glycogen following glycogen-depleting exercise(Reference Casey, Mann and Banister5). Furthermore, recent 13C-MRS studies reveal that ingestion of MD with either fructose or galactose (GAL) are twice as effective at restoring liver glycogen compared with MD plus glucose(Reference Decombaz, Jentjens and Ith22). Fructose and GAL are primary substrates for the liver, as they are metabolised in the liver for release of glucose or stored as glycogen for subsequent release(Reference Williams23, Reference Nilsson and Hultman24), whereas glucose is not exclusively a direct substrate for liver glycogen synthesis(Reference Nilsson and Hultman24, Reference Newgard, Hirsch and Foster25). The use of GAL is more attractive than fructose, as GAL combined with MD has been shown to produce marginally greater increases in liver glycogen content in comparison to MD plus fructose (179 v. 154 mmol/l(Reference Decombaz, Jentjens and Ith22)). This suggests that it may support endurance performance more effectively, though this is yet to be established.
The purpose of the present study was to compare liver and muscle glycogen repletion following glycogen-depleting exercise when a fractional part of MD in an isoenergetic MD and GAL formulation was substituted with PRO and AA. The use of in vivo 13C-MRS made it possible to establish non-invasively the effects of these formulations on liver (adding to this relatively sparse literature) and muscle glycogen metabolism. To date, no studies have reported whether the beneficial effect of mixing CHO and PRO with AA upon glycogen synthesis in muscle post-exercise also applies to liver glycogen synthesis, especially as the role of insulin has been deemed important(Reference Roden, Perseghin and Petersen26). Further, the combined use of MD and GAL has been shown to be most effective at enhancing liver glycogen(Reference Decombaz, Jentjens and Ith22). Therefore, we hypothesised that the post-exercise ingestion of MD and GAL with PRO and AA would enhance liver and muscle glycogen repletion compared with an isoenergetic MD–GAL formulation.
Methods
Participants
A total of seven endurance-trained (approximatley 10 h per week, for at least 5 years) male cyclists (age: 33 (se 8) years; BM: 79 (se 9) kg; stature: 178 (se 7) cm; maximal oxygen uptake (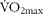 $\dot {>\%V}O_{2max}$): 58 (se 7) ml/kg per min) took part in the study. The study was conducted according to the guidelines laid down in the Declaration of Helsinki, and all procedures involving human participants were approved by the Leeds Metropolitan University ethics committee. Procedures and potential risks were explained to each participant prior to the study and written consent was obtained.
$\dot {>\%V}O_{2max}$): 58 (se 7) ml/kg per min) took part in the study. The study was conducted according to the guidelines laid down in the Declaration of Helsinki, and all procedures involving human participants were approved by the Leeds Metropolitan University ethics committee. Procedures and potential risks were explained to each participant prior to the study and written consent was obtained.
Preliminary testing
Participants completed a maximal incremental cycle test to volitional exhaustion to determine their individual $\dot {>\%V}O_{2max}$ and maximal workload (W max; mean for all participants: 350 (se 17) W), as described previously(Reference Kuipers, Keizer and Brouns27). This preceded the experimental trials by at least 1 week. Using regression analysis, relative oxygen uptake and power output were then used to determine the relative exercise intensities to be undertaken by each participant during the experimental trials (i.e. power output (W) at a given % $\dot {>\%V}O_{2max}$).
Diet and physical activity instructions
Participants were asked to record their food intake and refrain from any physical activity 48 h prior to the initial experimental trial. They were then instructed to follow the same diet and exercise activities before the second trial. Participants were required to refrain from the consumption of alcohol and caffeine for 24 h preceding each of the experimental trials. The night before each trial, participants were provided with a standardised meal (4312 kJ) containing 150 g CHO (55 %), 67 g PRO (26 %) and 22 g fat (19 %).
Experimental trials
Following a 10 h overnight fast, participants started their experimental trials at the same time of the day (between 06.00 and 09.00 hours). Upon arrival at the laboratory, a catheter was inserted into a cephalic vein for regular blood sampling. Baseline blood samples were drawn 5 min before exercise. All blood samples were analysed for plasma glucose, lactate, NEFA and serum insulin concentration, as described previously(Reference Trenell, Stevenson and Stockmann28).
Participants then completed a glycogen-depleting exercise challenge (adapted from Bowtell et al. (Reference Bowtell, Gelly and Jackman29), Fig. 1) on a cycle ergometer (SRM Training Systems). Participants cycled for 45 min at 70 $\dot {>\%V}O_{2max}$, before completing six 1 min sprints at 120 $\dot {>\%V}O_{2max}$ separated by a 2 min recovery at 50 $\dot {>\%V}O_{2max}$. Subsequently, participants cycled for another 45 min at 70 $\dot {>\%V}O_{2max}$ in order to further promote depletion of glycogen in type I fibres, as well as reduction of plasma lactate concentrations at the end of the glycogen-depleting exercise(Reference Vollestad and Blom30). Participants were allowed to consume water ad libitum during the exercise period and this then formed their prescription during the second experimental trial.

Fig. 1 Schematic representation of experimental procedures. 13C-MRS, 13C magnetic resonance spectroscopy; GI, gastrointestinal. ![]() , Warm-up at 50 % maximal oxygen uptake ($\dot {>\%V}O_{2max}$);
, Warm-up at 50 % maximal oxygen uptake ($\dot {>\%V}O_{2max}$); ![]() , 45 min at 70 % $\dot {>\%V}O_{2max}$;
, 45 min at 70 % $\dot {>\%V}O_{2max}$; ![]() , 6 × 1 min sprints at 120 %
, 6 × 1 min sprints at 120 %  $\dot {>\%V}O_{2max}$.
$\dot {>\%V}O_{2max}$.
Blood samples were drawn at the start, at 45 min into the 4 h recovery period and at every 30 min thereafter. Immediately after glycogen-depleting exercise and at the end of the 4 h recovery period, muscle (vastus lateralis) and liver glycogen concentrations were acquired using a 3T Achieva whole body scanner (Philips). Muscle spectra were acquired using a 13C/1H leg coil (PulseTeq) and liver spectra using an in-house built 13C/1H liver coil (13C coil diameters of 6 and 12 cm for leg muscle and liver, respectively).
Immediately following the initial 13C-MRS scan (45 min into the recovery period), participants ingested the first bolus (approximately 600 ml, 22 % of total volume adjusted for BM) of the MD (0·9 g/kg BM per h (Roquette)) and GAL (0·3 g/kg BM per h) beverage or the MD (0·5 g/kg BM per h), GAL (0·3 g/kg BM per h) and PRO+AA (0·4 g/kg BM per h (0·2 g/kg BM per h of whey PRO hydrolysate (ARLA Foods), 0·1 g/kg BM per h of l-leucine (Fagron) and 0·1 g/kg BM per h l-phenylalanine (Fagron))) beverage. The beverages were presented using a randomised, double-blind study design. Both beverages contained 0·58 g NaCl/l, as well as sweetener and flavouring. Subsequent boluses were presented at every 30 min thereafter (each approximately 350 ml, 13 % of total volume, 2·7 litres).
Questionnaires
Participants were monitored for symptoms of gastrointestinal distress using a ten-point Likert scale upon completion of the glycogen-depleting exercise and at every 30 min from 45 min into the 4 h recovery period.
Magnetic resonance spectroscopy
Skeletal muscle
With the participant supine, the leg coil was placed equidistant between the caudal tip of the patella and the caudal head of the femur over the vastus lateralis, and its position was marked with indelible ink. Scout images were acquired and the distance between coil surface and the muscle was recorded to ensure identical coil positioning on repeat 13C scans. The 13C pulse power was calibrated to a nominal value of 80° in the tissue of interest by observing the power-dependent variation in signal from a fiducial marker located in the coil housing, containing a sample exhibiting a 13C signal with short T1 (50 % acetone, 50 % water, 25 mm-GdCl3). Spectra showing the glycogen-C1 natural abundance 13C resonance were acquired using a non-localised 1H-decoupled 13C pulse-acquire sequence (relaxation time = 230 ms, spectral width = 8 kHz, 3000 averages, WALTZ decoupling, nominal tip angle = 80°) over a 12 min acquisition time.
Liver
The participant was placed in a supine position and the liver coil was placed centrally over the liver, confirmed by scout images. The distance between the coil surface and the liver was noted. The 13C pulse power was calibrated to a nominal value of 80° in the tissue of interest by observing the power-dependent variation in signal from a fiducial marker, as described for the skeletal muscle methodology. Spectra showing the glycogen-C1 13C resonance were acquired using a non-localised 1H-decoupled 13C pulse-acquire sequence (relaxation time = 300 ms, spectral width = 8 kHz, 2000 averages, WALTZ decoupling, nominal tip angle = 80°) over a 12 min acquisition time.
Analysis and quantitation of spectra
All spectra were analysed using ‘Java-based magnetic resonance user interface’ software (jMRUI version 3·0; http://www.mrui.uab.es/mrui/) and the AMARES algorithm(Reference Naressi, Couturier and Castang31, Reference Naressi, Couturier and Devos32). Glycogen content was determined from the magnitude of natural abundance C1-glycogen signal at 100·5 parts per million (ppm). Quantification of 13C spectra was achieved by comparison of the in vivo glycogen-C1 13C signal amplitudes with that of a standard glycogen solution (100 mm-glycogen, 70 mm-KCl, 0·05 % sodium azide). Quantitated 13C spectra were acquired from leg- and liver-shaped phantoms at a range of separations between coil and phantom, to account for the separation between coil and muscle or liver due to skin, subcutaneous fat and/or the rib cage, using the same coils, pulse sequences and tip angles as employed for in vivo spectra.
Statistical analysis
Statistical analyses were performed using SPSS for Windows (version 17.0, SPSS, Inc.). The normal distribution of data was assessed by the use of the Kolmogorov–Smirnov test. A two-way ANOVA for repeated measures was employed to compare differences in plasma glucose, plasma lactate, plasma NEFA and serum insulin concentrations (time and treatment). Where significance was detected, post hoc analysis was performed using a paired t test with Bonferroni adjustment. A two-way ANOVA was also used to compare differences in absolute muscle and liver glycogen concentrations over time and between conditions. Where a main effect of time was detected, post hoc analysis was performed using a paired t test with Bonferroni adjustment. The rates of muscle and liver glycogen repletion were analysed using a related samples t test. Ratings of gastrointestinal distress were analysed by the Friedman rank test with the Wilcoxon signed rank test. A level of confidence denoting statistical difference was set at 0·95 (P< 0·05), with actual P values reported. All the values are means with their standard errors unless otherwise stated.
Results
Liver and muscle glycogen concentrations
There were no significant differences between trials for liver and muscle glycogen concentrations following glycogen-depleting exercise (135·3 (se 21·7) and 26·7 (se 3·5) mmol/l for MD–GAL; 161·7 (se 15·8) and 36·4 (se 7·3) mmol/l for MD–GAL-PRO +AA, respectively (Fig. 2)). Post-recovery glycogen concentrations were significantly higher than following depletion exercise for both liver (195·3 (se 22·6) (P= 0·01) v. 213·8 (se 18·0) mmol/l (P= 0·001)) and muscle (49·7 (se 4·0) (P= 0·000) v. 51·1 (se 7·9) mmol/l (P= 0·001)) for MD–GAL and MD–GAL-PRO+AA, respectively (Fig. 2). There was no significant difference between treatment conditions.

Fig. 2 Individual (a, b) liver and (c, d) intramuscular glycogen concentrations after glycogen-depleting exercise and following the recovery period for the (a, c) maltodextrin (MD)-galactose (GAL) and (b, d) MD–GAL-protein+amino acids treatments (n 7). ![]() , Mean for each treatment.
, Mean for each treatment.
The rate of liver glycogen repletion (Fig. 3(a)) over the recovery period was not significantly different between trials (15·0 (se 2·5) v. 13·0 (se 2·7) mmol/l per h for MD–GAL and MD–GAL-PRO+AA, respectively). The rate of muscle glycogen repletion (Fig. 3(b)) was significantly (P= 0·04) higher following MD–GAL (5·8 (se 0·7) mmol/l per h) in comparison to MD–GAL-PRO+AA (3·7 (se 0·6) mmol/l per h).

Fig. 3 Individual rates of (a) liver and (b) intramuscular glycogen repletion for the recovery period for the maltodextrin (MD)-galactose (GAL) and MD–GAL-protein (PRO)+amino acids (AA) treatments (n 7). ![]() , Mean for each treatment. * MD–GAL significantly higher than MD–GAL-PRO+AA (P< 0·05).
, Mean for each treatment. * MD–GAL significantly higher than MD–GAL-PRO+AA (P< 0·05).
Blood metabolites
Fasting plasma glucose concentrations were not significantly different between trials (average 4·7 (se 0·1) mmol/l). Plasma glucose, serum insulin and plasma NEFA concentrations during recovery are shown in Fig. 4(a), (b) and (c), respectively. Plasma glucose concentrations increased during recovery following the initial bolus of both treatments, with MD–GAL reaching a significantly (P= 0·003) higher peak at 75 min (7·8 (se 0·5) mmol/l (30 min after the initial bolus)) in comparison to MD–GAL-PRO+AA (6·0 (se 0·2) mmol/l). Plasma glucose concentrations were significantly (P= 0·001–0·015) higher for the majority of the remaining recovery period during MD–GAL in comparison to MD–GAL-PRO+AA.

Fig. 4 Mean (a) plasma glucose, (b) serum insulin, (c) NEFA and (d) plasma lactate during the recovery period for the maltodextrin (MD)-galactose (GAL) (![]() ) and MD–GAL-protein (PRO)+amino acids (AA) (
) and MD–GAL-protein (PRO)+amino acids (AA) (![]() ) treatments. Values are means (n 7), with their standard errors represented by vertical bars. * MD–GAL significantly higher than MD–GAL-PRO+AA (P< 0·05). MRS, magnetic resonance spectroscopy. To convert insulin from mu to pmol, multiply by 6·945.
) treatments. Values are means (n 7), with their standard errors represented by vertical bars. * MD–GAL significantly higher than MD–GAL-PRO+AA (P< 0·05). MRS, magnetic resonance spectroscopy. To convert insulin from mu to pmol, multiply by 6·945.
Fasting serum insulin concentrations were not significantly different between MD–GAL and MD–GAL-PRO+AA (3·8 (se 0·8) v. 4·3 (se 0·5) mU/l; 26·4 (se 5·6) v. 29·9 (se 3·5) pmol/l). Following glycogen-depleting exercise and the initial bolus of both treatments, serum insulin concentrations increased significantly (P= 0·03) with MD–GAL peaking earlier (195 min (150 min after the initial bolus)) than MD-GAL-PRO+AA (225 min (180 min after the initial bolus)). The MD–GAL-PRO+AA had significantly higher peak insulin compared with MD–GAL (97·2 (se 12·6) v. 43·9 (se 8·9) mU/l, (675·1 (se 87·5) v. 304·9 (se 61·8) pmol/l) P= 0·01). Peak insulin concentrations declined to 18·6 and 22·6 mU/l (129·2 and 157·0 pmol/l) for MD–GAL and MD–GAL-PRO+AA, respectively. From 75 min (30 min after the initial bolus) into the recovery MD–GAL-PRO+AA showed significantly (P= 0·005–0·033) higher serum insulin concentrations for the majority of the remaining recovery period.
Fasting plasma NEFA concentrations were not significantly different between MD–GAL and MD–GAL-PRO+AA trials (Fig. 4(c)). There was no significant interaction between treatments at any time point. Plasma lactate concentrations were elevated in the MD–GAL-PRO+AA from 195 min (150 min after the initial bolus) onwards in comparison to the MD–GAL (Fig. 4(d), P= 0·000–0·005).
Gastrointestinal distress
The median scores for gastrointestinal distress were higher at 3·5 and 4 h of recovery for MD–GAL-PRO+AA (4 and 5, respectively) compared with a score of 1 during the preceding recovery period, but there was no time or time and condition interaction.
Discussion
The present study is the first to evaluate, using 13C-MRS, the effects of MD and GAL, with and without PRO and AA, on liver and muscle glycogen synthesis, during a 4 h recovery from glycogen-depleting exercise. The primary findings of the present study are that, despite the greater postprandial insulin response, the ingestion of MD and GAL with PRO and AA did not enhance, but matched, the final liver and muscle glycogen repletion compared with an isoenergetic MD–GAL mix. The value of glycogen reserves in muscle and liver is related to endurance performance(Reference Casey, Mann and Banister5, Reference Bergstrom, Hermansen and Hultman6), but the consumption of PRO and AA has other potential benefits, such as post-exercise muscle PRO synthesis. This can positively affect gains in muscle PRO and strength(Reference Levenhagen, Gresham and Carlson33), which to an athlete would be worth considering as part of any recovery strategy.
The relative increase in liver glycogen concentration for the MD–GAL (52 %) and MD–GAL-PRO+AA (35 %) trials represent mean rates of recovery of 15 and 13 mmol/l per h, respectively. The rates of recovery are lower than previous studies(Reference Decombaz, Jentjens and Ith22), which have shown that the combination of MD and GAL (28 mmol/l per h) is twice as effective as MD and glucose. However, the amount of MD and GAL (approximately 380 g v. approximately 530 g) and duration of recovery (4 v. 6·5 h) may be an explanation for the differences between these reports. In addition, the lower post-depletion liver glycogen concentrations in the previous report, compared with the present study (82 v. 135 mmol/l), may be influential, as liver glycogen synthesis has been suggested to be inversely related to liver glycogen concentration(Reference Fleig, Enderle and Steudter34). Nevertheless, the post-depletion glycogen concentrations in the present study are comparable with other reports (141–191 mmol/l(Reference Casey, Mann and Banister5)) and show marked depletion compared with typical fasting concentrations (approximately 250 to approximately 360 mmol/l(Reference Petersen, Price and Bergeron3, Reference Roden, Perseghin and Petersen26)).
Insulin plays an important role in liver glycogen synthesis, with portal vein insulin concentrations of approximately 19–24 mU/l (130–170 pmol/l) signalling half-maximal stimulation of glycogen synthesis(Reference Roden, Perseghin and Petersen26). Therefore, the present study included insulinogenic AA within one of the trials to establish whether their addition would further enhance liver glycogen synthesis from MD. These data show that the partial replacement of MD with PRO and AA in a MD–GAL mix initiated a greater postprandial insulin response and did not enhance, but matched, the overall rate of liver glycogen repletion compared with MD–GAL.
The similarity in liver glycogen repletion between the trials may be explained by portal vein insulin concentrations, which are known to be approximately three times higher than peripheral concentration(Reference Horwitz, Starr and Mako35) (estimated in this case to be 90 and 184 mU/l (625 and 1278 pmol/l) for MD–GAL and MD–GAL-PRO+AA, respectively, calculated from peripheral mean serum insulin concentrations during the recovery period of 30 and 61 mU/l (208 and 424 pmol/l)). It is possible that, even without the addition of PRO and AA, portal vein insulin concentrations were sufficient for maximal glycogen repletion and that the PRO-AA-induced hyperinsulinaemia conferred no additional benefit. Therefore, the facts that the addition of PRO and AA was made on the basis of a direct isoenergetic substitution for MD and that final liver glycogen concentrations were similar would suggest that some compensation for the lower MD content was achieved. It is likely that GAL provided a significant source of substrate for liver glycogen repletion and the contribution from MD may have been minimal. However, there is the possibility that the addition of GAL to MD is likely to further increase the activation of glycogen synthase and decrease glycogen phosphorylase activity(Reference Sparks, Lynch and Glinsmann36). Thus, GAL may facilitate a significant contribution to liver glycogen resynthesis from MD. These observations suggest that there is no disadvantage in substituting some of the MD in an MD–GAL mix with PRO and AA for effective liver glycogen repletion to improve subsequent exercise performance.
The substitution by whey PRO and insulinogenic AA (leucine and phenylalanine) to a fractional part of MD in a MD–GAL mix produced a doubling of the postprandial insulin response. The insulin response is comparable with that reported by van Loon et al. (Reference van Loon, Saris and Kruijshoop8, Reference van Loon, Saris and Verhagen15), who used a similar combination of PRO and AA. It was only when MD–GAL was consumed with PRO and AA that insulin concentrations exceeded the minimum proposed (approximately 47 mU/l; 326 pmol/l) necessary to promote glucose storage(Reference Young, Bogardus and Stone37). These present data demonstrate that elevating circulatory insulin levels with reduced MD availability did not enhance, but matched, absolute muscle glycogen storage, again suggesting a compensatory effect of PRO and AA for a lower amount of MD. However, the long-term health consequences of such hyperinsulinaemia need to be established and should be taken into practical consideration.
The muscle glycogen repletion rate of 5·8 mmol/l per h, estimated to be 5·2 mmol/kg WW per h (assuming skeletal muscle mass density of 1·1 g/cm3(Reference Ward and Lieber38)) in the present study following MD (0·9 g/kg BM per h) and GAL (0·3 g/kg BM per h), is comparable with rates of 3·9–10·5 mmol/kg WW per h reported for doses of CHO between 0·8 and 1·2 g/kg BM per h(Reference van Loon, Saris and Kruijshoop8, Reference Jentjens, van Loon and Mann10, Reference Zawadzki, Yaspelkis and Ivy14, Reference Decombaz, Jentjens and Ith22). The partial replacement of MD with PRO and AA statistically reduced muscle glycogen repletion rates, despite the higher postprandial insulin response compared with an isoenergetic amount of MD–GAL. This suggests that MD availability is a limiting factor for muscle glycogen repletion. A greater advantage could be seen if PRO and AA were added to MD (0·9 g/kg BM per h) and GAL (0·3 g/kg BM per h), which is yet to be established.
The lower rate of muscle glycogen repletion for MD–GAL-PRO+AA is unlikely to be related to differences in gastric emptying between the treatments, as insulin and glucose concentrations rose rapidly following the ingestion of both treatments. The plasma glucose response was attenuated during recovery following MD–GAL-PRO+AA, which is a possible reflection of a reduced rate of glucose appearance in the circulation rather than glucose disposal(Reference Kaastra, Manders and Van Breda39). However, this simply may be due to the reduced amount of MD in this treatment, not resulting in a greater glycaemic response compared with MD–GAL and thereby limiting the rate of glycogen repletion. Furthermore, any differences in gastric emptying did not seem to affect liver glycogen repletion rates. It is possible that a proportion of the feedings may have remained in the gastrointestinal space towards the end of the recovery period (3·5 and 4 h), as gastrointestinal distress started to increase for MD–GAL-PRO+AA. It is therefore possible that a longer interval between the ingestion of the last bolus and post-recovery scan may have resulted in a higher rate of muscle glycogen in the MD–GAL-PRO+AA trial.
Muscle glycogen concentrations post-exercise were depleted in comparison to typical fasting concentrations (95–150 mmol/l(Reference Stevenson, Thelwall and Thomas4, Reference Berardi, Price and Noreen16, Reference Ivy, Goforth and Damon17)) and are comparable with the literature (approximately 40–55 mmol/l(Reference Stevenson, Thelwall and Thomas4, Reference Berardi, Price and Noreen16, Reference Ivy, Goforth and Damon17)). Muscle glycogen concentrations were lower for MD–GAL in comparison to MD–GAL-PRO+AA (27 % difference), though not significantly, which is comparable with the literature(Reference Casey, Mann and Banister5, Reference Betts, Williams and Boobis19). Lower glycogen concentrations have an inverse relationship with glycogen synthesis, as long as muscle glycogen concentrations are below the range (28–65 mmol/kg WW) that has been shown to have no effect(Reference Price, Rothman and Taylor13). The mean muscle glycogen concentrations for MD–GAL were 27 mmol/l, estimated to be 24 mmol/kg WW(Reference Ward and Lieber38) in the present study. However, there was no consistent response for individuals having muscle glycogen concentrations equivalent to < 28 mmol/kg WW during MD–GAL and ≥ 28 mmol/kg WW during the MD–GAL-PRO+AA treatment. Therefore, this difference is unlikely to have affected muscle glycogen repletion rates. Muscle glycogen repletion can occur via indirect pathways, especially when there is high precursor availability(Reference Nordheim and Vollestad40). Therefore, the higher plasma lactate concentrations during the latter part of the 4 h recovery period during MD–GAL may explain, in part, its higher muscle glycogen repletion rate compared with MD–GAL-PRO+AA.
The delayed provision of both treatments until after the completion of the liver glycogen measurement (approximately 45 min post-depletion exercise), to prevent contamination of the measurement(Reference Decombaz, Jentjens and Ith22), may have affected the ability of PRO to increase muscle glycogen repletion. PRO and CHO ingested have been shown to positively effect the insulin-independent phase of glycogen recovery(Reference Ivy, Goforth and Damon17). Previous reports observed a 2-fold increase in muscle glycogen storage rates during the initial 40 min of recovery, with a CHO PRO treatment compared with an isoenergetic CHO treatment(Reference Ivy, Goforth and Damon17). Therefore, it is possible that if both treatments were provided earlier in the insulin-independent phase following exercise, muscle glycogen recovery per se may have been greater, especially for the MD–GAL-PRO+AA treatment. There is also the possibility that the inclusion of GAL may have decreased the availability of MD for muscle glycogen repletion. GAL has been shown in vitro to activate liver glycogen synthase activity(Reference Sparks, Lynch and Glinsmann36), meaning that the liver and muscle may have been in direct competition for the available glucose from MD.
In conclusion, despite the elevated insulin response with the addition of PRO and AA in the MD–GAL mix compared with an isoenergetic mix of MD–GAL, this did not confer additional benefits for the short-term restoration of absolute liver and muscle glycogen concentrations following glycogen-depleting exercise. However, we have established that the elevated insulin response, with the addition of PRO and AA in the MD–GAL mix, compared with MD–GAL, compensated for the lower MD content, producing similar rates and repletion of liver glycogen and, in the case of muscle, to a similar final glycogen concentration. Therefore, the provision of PRO and AA may be beneficial during scenarios when CHO availability, or time, is restricted between bouts of exercise.
Acknowledgements
The authors thank all the volunteers for taking part in the study. They also thank Ben Taylor and Laurie Patterson for their assistance with data acquisition and Carol Smith, Louise Morris and Tim Hodgson for their assistance with 13C-MRS data acquisition. This work was supported by the Yorkshire Concept, UK and the MoveLab, UK. All authors significantly contributed to the study and read and approved the final content of the manuscript. E. D. contributed to the study design, data acquisition and analysis and was the main contributor to the writing of the manuscript. J. P. O'H. contributed to the study design, data acquisition and discussion, and was the second main contributor to the writing of the manuscript. P. E. T. contributed to the study design, data acquisition and analysis and the writing of the manuscript. F. E. S. contributed to the study design, data acquisition and analysis. D. G. J. contributed to the data acquisition and discussion. R. F. G. J. K. contributed to the study design, analysis, discussion and the writing of the manuscript. M. I. T. contributed to the study design, data acquisition, analysis, discussion and the writing of the manuscript. The authors declare no conflict of interest.




