
Contents
Research Article
Twins and Chronogenetics: Correlated Pathways of Development
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 149-157
-
- Article
-
- You have access
- Export citation
Twin Development and Temperament
Within-Pair Differences in Newborn Twins: Effects of Gender and Gestational Age on Behavior
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 159-164
-
- Article
-
- You have access
- Export citation
Features of Interaction in Infant Twins
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 165-171
-
- Article
-
- You have access
- Export citation
Childhood Sex-Role Behaviors: Similarities and Differences in Twins
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 173-179
-
- Article
-
- You have access
- Export citation
Twin Similarity in the Developmental Transformations of Infant Temperament as Measured in a Multi-Method, Longitudinal Study
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 181-189
-
- Article
-
- You have access
- Export citation
The Role of Parental Attitudes in the Development of Temperament in Twins at Home, School and in Test Situations
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 191-204
-
- Article
-
- You have access
- Export citation
Self- and Teacher-Rated School Adjustment in MZ and DZ Twins
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 205-212
-
- Article
-
- You have access
- Export citation
Twins and Mental Abilities
Similarities in Test Scores and Profiles of Kibbutz Twins and Singletons
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 213-218
-
- Article
-
- You have access
- Export citation
Does a Special Twin Situation Contribute to Similarity for Abilities in MZ and DZ Twins?
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 219-222
-
- Article
-
- You have access
- Export citation
The High Incidence of Reading Disability in Twin Boys and Its Implications for Genetic Analyses
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 223-236
-
- Article
-
- You have access
- Export citation
Blood Uric Acid Level and IQ: A Study in Twin Families
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 237-242
-
- Article
-
- You have access
- Export citation
Twins Reared Apart
Swedish Early Separated Twins: Identification and Characterization
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 243-250
-
- Article
-
- You have access
- Export citation
Finnish Twins Reared Apart II: Validation of Zygosity, Environmental Dissimilarity and Weight and Height
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 251-258
-
- Article
-
- You have access
- Export citation
-
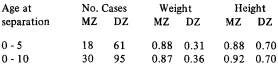
Within the Finnish Twin Cohort of like-sexed adult twin pairs, a subgroup of pairs separated at an early age has been identified. In 165 pairs, both cotwins responded to questionnaires in 1975 and 1979. An environmental dissimilarity score was formed which consists of items on whether the twins had lived after separation in the same community, attended the same school, were on the same grade at school, how often the cotwins met, how often they met common friends and relatives and whether they attended the same clubs etc, or not. To validate the zygosity diagnosis obtained by questionnaire in 1975, those pairs whose zygosity was unknown as well as those with the least contact after separation were contacted for blood sampling (11 bloodgroups). Of 15 pairs with no zygosity diagnosis, 10 responded (1 no address,2 abroad,2 refused). Six pairs were classified MZ and 4 DZ. In 12 MZ and 8 DZ pairs undergoing bloodgroup determination, the classification of only one pair changed from DZ to MZ. The following intraclass correlations for height and weight were found:

Finnish Twins Reared Apart III: Personality Factors
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 259-264
-
- Article
-
- You have access
- Export citation
-
This study is based on data from 165 adult twin pairs separated at 10 years or less. Information on personality factors: extraversion (E) and neuroticism (N) (EPI scale short from), life satisfaction (LS) (Allardt) and stress of daily activities (SDA) was obtained as part of the questionnaire study carried out in the entire Finnish Twin Cohort in 1975. Later in 1979 a questionnaire sent to the twins reared apart yielded a scale (range 7-30 points) measuring the environmental dissimilarities after separation (reliability 0.83). The effect of separation on personality factors by analysis of variance of individual data was studied. Sex, zygosity and age-at-separation were included in the models. The overall expalanatory rates were low (2.1-4.4%). The definitive study group was formed by selecting those pairs with a dissimilarity score greater than 15. The following intraclass correlations were obtained:


Behavior Genetic Analysis
Altruism and Genetics
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 265-271
-
- Article
-
- You have access
- Export citation
A Bivariate Path Analysis of Fear Data on Twins and Their Parents
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 273-286
-
- Article
-
- You have access
- Export citation
Detecting Sex-Associated Variation With the Families of MZ and DZ Twins
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 287-301
-
- Article
-
- You have access
- Export citation
Genetics of Psychopathology
Behavioural Genetics of Early Childhood: Fears, Restlessness, Motion Sickness and Enuresis
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 303-306
-
- Article
-
- You have access
- Export citation
Health Related Psychosocial Correlates of Neuroticism: A Study of Adult Male Twins in Finland
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 307-320
-
- Article
-
- You have access
- Export citation
Psychiatric Hospitalization in Twins
-
- Published online by Cambridge University Press:
- 01 August 2014, pp. 321-332
-
- Article
-
- You have access
- Export citation