



For nearly 2,000 years, people in the US Southwest and Northwest Mexico (SW/NW) have woven the turkey (Meleagris gallopavo) into aspects of their economic, social, and ceremonial life (Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016, Reference Lipe, Tushingham, Blinman, Webster, LaRue, Oliver-Bozeman and Till2020; McKusick Reference McKusick1986, Reference McKusick2001; McNeil Reference McNeil2022; Munro Reference Munro and Smith2011). For example, some prehispanic groups relied on this bird for protein (Badenhorst and Driver Reference Badenhorst and Driver2009; Ellyson et al. Reference Ellyson, Nagaoka and Wolverton2019), and they used the feathers and bones for clothing, ritual paraphernalia, tools, and musical instruments (Blinman Reference Blinman2001; Lipe et al. Reference Lipe, Tushingham, Blinman, Webster, LaRue, Oliver-Bozeman and Till2020; Payne Reference Payne1991). People also depicted turkeys on rock art and pottery, they buried these birds in ritual contexts (Hill Reference Hill2000), and turkeys are central figures in oral traditions (McNeil Reference McNeil2022; Tyler Reference Tyler1979:85–106). Turkeys and dogs were the only domesticated animals people had in this region before European arrival, so understanding the cultural and environmental context of how, why, when, and where people managed turkeys contributes to issues in anthropological archaeology and social zooarchaeology (Kintigh et al. Reference Kintigh, Altschul, Beaudry, Drennan, Kinzig, Kohler and Fredrick Limp2014:17; Russell Reference Russell2012).
Archaeologists study turkeys in many ways, including visual and osteometric analyses of bones to infer sex, age, and trauma (Badenhorst et al. Reference Badenhorst, Lyle, Merewether, Driver and Ryan2012; Fothergill Reference Fothergill2016; McKusick Reference McKusick1986; Senior and Pierce Reference Senior and Pierce1989), and we use scanning electron microscopy on eggshells to understand their biological development (Beacham and Durand Reference Beacham and Durand2007; Conrad et al. Reference Conrad, Jones, Newsome and Schwartz2016). However, the two most popular methods of studying turkeys recently are ancient mitochondrial DNA (mtDNA) analysis to determine maternal genetic lineages (Kemp et al. Reference Kemp, Judd, Monroe, Eerkens, Hilldorfer, Cordray, Schad, Reams, Ortman and Kohler2017; Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016; Manin et al. Reference Manin, Corona-M, Alexander, Craig, Thornton, Yang, Richards and Speller2018; Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010), and stable carbon (δ13C) and nitrogen (δ15N) isotope analyses to infer diet (Conrad et al. Reference Conrad, Jones, Newsome and Schwartz2016; Jones et al. Reference Jones, Conrad, Newsome, Kemp and Kocer2016; McCaffery et al. Reference McCaffery, Tykot, Gore and DeBoer2014, Reference McCaffery, Miller and Tykot2021; Rawlings and Driver Reference Rawlings and Driver2010).
Researchers using mtDNA and stable isotopes have made considerable advances in establishing how people in parts of the SW/NW managed turkeys. For example, prehispanic aviculturists—the people who raised and managed birds—fed turkeys maize, and they predominantly kept turkeys belonging to the extinct domesticated clade, which researchers have termed haplogroup H1 (Kemp et al. Reference Kemp, Judd, Monroe, Eerkens, Hilldorfer, Cordray, Schad, Reams, Ortman and Kohler2017; Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016; McCaffery et al. Reference McCaffery, Tykot, Gore and DeBoer2014, Reference McCaffery, Miller and Tykot2021; Rawlings and Driver Reference Rawlings and Driver2010; Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010). However, much of these data derive from the Ancestral Pueblo cultural regions of the northern Southwest. Less is known about the maternal genetic lineage, diet, and the ways people managed turkeys further south, including in the Mimbres Valley of southwestern New Mexico (Figure 1). To investigate how turkeys fit within Classic period Mimbres society from AD 1000 to 1130 (all dates are AD)—including subsistence, economic production, sociopolitical organization, and religious and ritual practice—we sampled 31 turkeys from Elk Ridge (LA 78963), Lake Roberts (LA 47821), Mattocks (LA 676), Old Town (LA 1113), and Wheaton-Smith (LA 18903) for mtDNA and stable isotope analyses. We focus on turkeys from the Classic because this period is marked by increasing social complexity with population growth, a heightened reliance on maize agriculture, transformations in architecture and pottery styles, and the emergence of distinct social and religious traditions (Blake et al. Reference Blake, LeBlanc and Minnis1986; Hegmon Reference Hegmon2002; LeBlanc Reference LeBlanc1983; Minnis Reference Minnis1985; Shafer Reference Shafer, Powell-Martí and Gilman2006). Also, as shown in Figure 2, Classic period potters painted turkeys on some of their vessels.
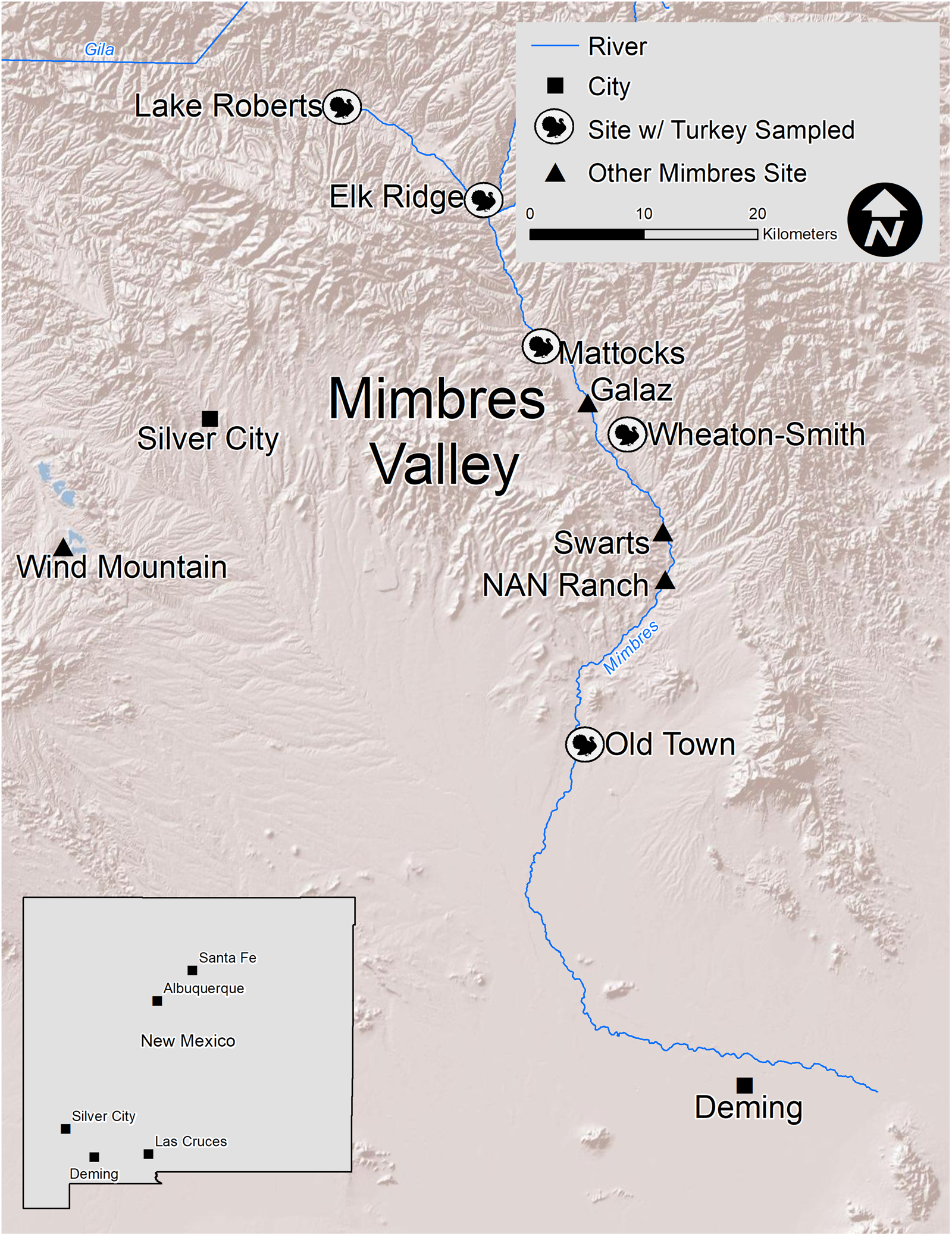
Figure 1. The Mimbres Valley in southwestern New Mexico, sites from which turkey remains were analyzed, and other sites mentioned in the text. (Map by Sean Dolan.)
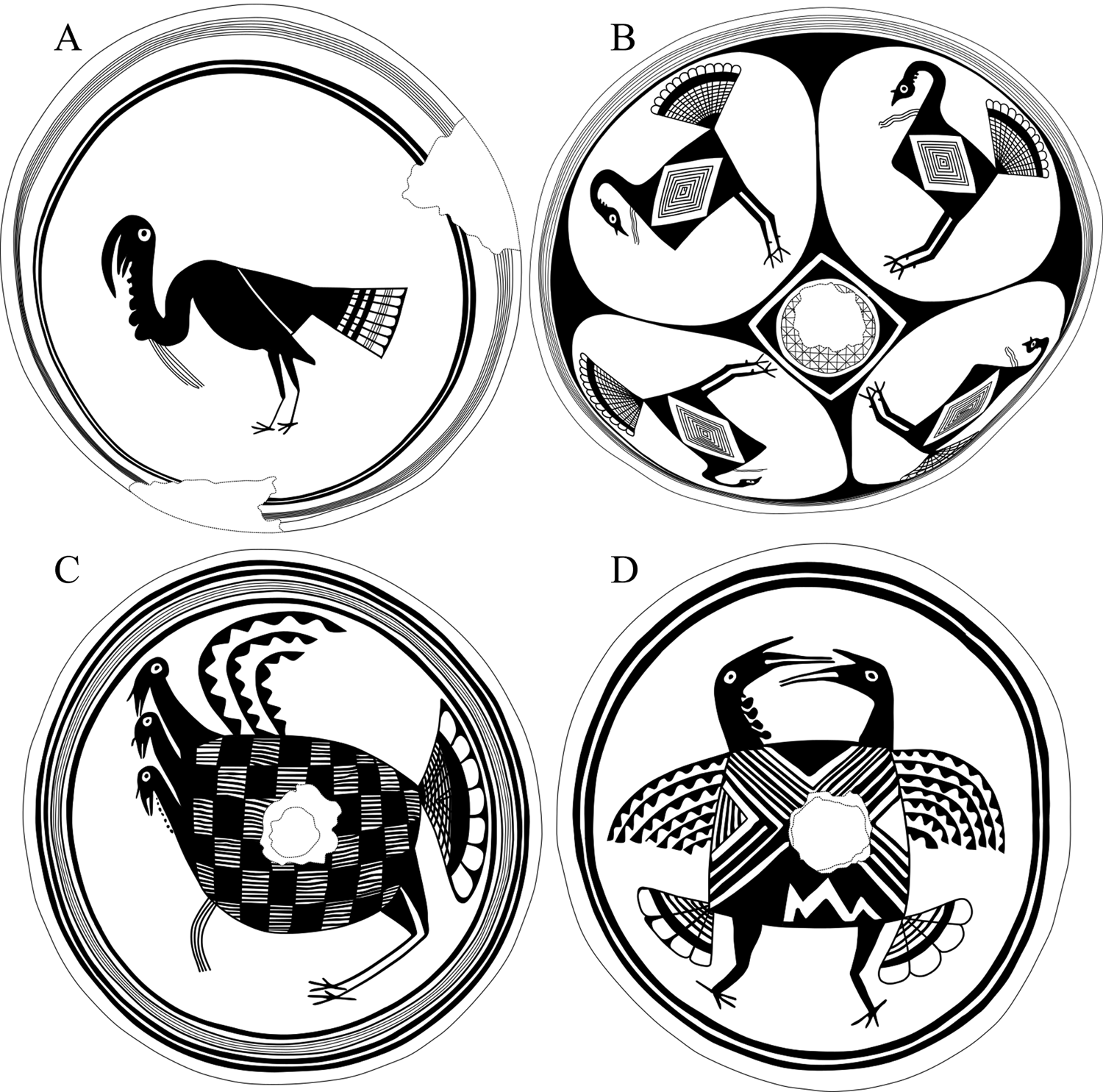
Figure 2. Examples of Mimbres pottery vessels that depict turkeys: (A) MA2222, from Swarts (after Cosgrove and Cosgrove Reference Cosgrove and Burton Cosgrove1932:Plate 216b); (B) MA9560, from Swarts (after Cosgrove and Cosgrove Reference Cosgrove and Burton Cosgrove1932:Plate 216d); (C) MA140, provenience unknown (after Fewkes Reference Fewkes1923:Figure 58); (D) MA5967, provenience unknown. (Vessel illustrations by Caitlin Wichlacz.)
This study examines the evidence for Classic period turkey management, diet, and genetic lineage in the Mimbres Valley. We begin with a brief discussion of what researchers have already published on turkey genetics and diet from the SW/NW, followed by an introduction to Classic period Mimbres society and what is known about turkeys from this region. We then introduce the five sites, sampled turkeys, and the methods used to determine the maternal genetic lineage and diet (in Supplemental Text 1–3), followed by the results. Results indicate that Mimbres aviculturists managed haplogroup H1 and H2 turkeys, and they used more haplogroup H2 turkeys than people in the northern Southwest. The Mimbres turkeys primarily consumed maize, but some had a nonmaize diet, and even fewer had a mixed diet. Moreover, although there is limited archaeological evidence for turkey pens in the Mimbres Valley, people limited turkey mobility. Also, unlike in parts of the northern Southwest, people in the Mimbres Valley did not eat turkeys (Morrison Reference Morrison and Beckett2002; Shaffer Reference Shaffer1991), and feather production outweighed any gains from their use as food, even during the Classic period, when turkey meat and fat would have helped combat environmental deficiencies.
Previous Turkey Studies
Using mtDNA analysis on turkey bones and coprolites, Speller and colleagues (Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010) found that SW/NW turkeys belong to two distinct maternal genetic lineages: haplogroups H1 and H2 (see also Jones et al. Reference Jones, Conrad, Newsome, Kemp and Kocer2016; Kemp et al. Reference Kemp, Judd, Monroe, Eerkens, Hilldorfer, Cordray, Schad, Reams, Ortman and Kohler2017; Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016). The H1 haplogroup is the most common in the SW/NW archaeological record, and it is virtually absent from the local wild subspecies, Merriam's wild turkey (Meleagris gallopavo merriami). This has led researchers to define turkeys of the H1 lineage as an extinct domesticated clade (Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016; Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010). The H1 haplogroup has a low genetic diversity, mainly represented by haplotype aHap1, highlighting over 1,600 years of breeding isolation from Merriam's turkeys (Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:98; Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010). The H2 haplogroup, however, derives from the Merriam's and Gould's wild turkey (Meleagris gallopavo mexicana) lineages (Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010). The H2 haplogroup archaeological specimens have genetic sequences similar to today's wild populations. Uncommon in the archaeological record, the H2 haplogroup is not the progenitor of the H1 lineage. Instead, the progenitor is related to the Rio Grande (Meleagris gallopavo intermedia) and Eastern (Meleagris gallopavo silvestris) subspecies that may have entered the region from the eastern United States in antiquity (Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010:2809).
Using the ratios of stable carbon (13C/12C) and nitrogen (15N/14N) isotopes in bone and teeth as biomarkers, researchers can infer the diet of ancient human and animal populations to answer archaeological, environmental, and evolutionary questions (Katzenberg Reference Katzenberg, Anne Katzenberg and Sanders2008). The proportion of stable carbon isotopes helps determine whether the animal ate a diet of temperate grasses, trees, and shrubs that have a C3 isotopic signature or a diet of tropical and savanna grasses—such as maize (Zea mays)—that have a C4 signature (Tykot Reference Tykot, Staller, Tykot and Benz2006). Most wild plants in the SW/NW have a C3 signature, but there are wild plants, such as amaranth (Amaranthus), that yield C4 signatures, and there are arid-adapted succulents, such as prickly pear (Opuntia sp.), that use a Crassulacean Acid Metabolism (CAM) photosynthetic pathway and that have δ13C values in between C3 and C4 plants (Tykot Reference Tykot, Staller, Tykot and Benz2006:132–133). Researchers use the proportion of stable nitrogen isotopes to understand the extent to which turkeys consumed protein from plant and animal sources (Rawlings and Driver Reference Rawlings and Driver2010:2435). However, absolute δ15N values may not be helpful given that δ15N values vary depending on the trophic level and dietary factors such as meat/vegetable ratio and marine versus terrestrial food sources (Ambrose Reference Ambrose1991; Schoeninger and DeNiro Reference Schoeninger and DeNiro1984).
Several studies focusing on human and turkey diets from the northern Southwest have identified high C4 maize consumption (Coltrain and Janetski Reference Coltrain and Janetski2013; Coltrain et al. Reference Coltrain, Janetski and Carlyle2007; Conrad et al. Reference Conrad, Jones, Newsome and Schwartz2016; Kellner et al. Reference Kellner, Schoeninger, Spielmann, Moore and Morgan2010; Martin Reference Martin1999; McCaffery et al. Reference McCaffery, Tykot, Gore and DeBoer2014; McCaffery et al. Reference McCaffery, Miller and Tykot2021; Rawlings and Driver Reference Rawlings and Driver2010). However, some turkeys consumed a C3 nonmaize diet, including a select few haplogroup H1 turkeys (Jones et al. Reference Jones, Conrad, Newsome, Kemp and Kocer2016). In addition, Lipe and colleagues (Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016) demonstrate that many haplogroup H2 turkeys from the northern Southwest consumed C4 diets. Consequently, as Jones and colleagues (Reference Jones, Conrad, Newsome, Kemp and Kocer2016:581) point out, “δ13C data cannot be used as a proxy for a turkey's mitochondrial DNA lineage, at least among Southwestern turkeys, without significant additional data.” As a result, we integrate both methods on individual turkeys from the Mimbres Valley to best understand management practices.
Classic Period Mimbres Society
The Mimbres Valley in southwestern New Mexico is the core of Mimbres culture, but the Mimbres region extends into other parts of New Mexico, Arizona, and northern Mexico. The Mimbres region is part of the larger Mogollon archaeological tradition that Haury (Reference Haury1936) first defined, and the cultural sequence is well established after a century of excavation, survey, and material culture studies (Anyon et al. Reference Anyon, Creel, Gilman, LeBlanc, Miller, Nash and Nelson2017). Archaeologists have published several summaries and edited volumes detailing the culture history, making the Mimbres Valley one of the most studied archaeological regions of the SW/NW (Hegmon Reference Hegmon2002; LeBlanc Reference LeBlanc1983; Lekson Reference Lekson2006; Powell-Martí and Gilman Reference Powell-Martí and Gilman2006; Roth et al. Reference Roth, Gilman and Anyon2018).
People in the Mimbres Valley lived in subsurface pithouse structures beginning approximately in the 200s, which is the same time they began making plain brown pottery. They transitioned to living in aboveground surface cobble masonry pueblos organized into room blocks around AD 1000 (Gilman Reference Gilman1987; Shafer Reference Shafer, Powell-Martí and Gilman2006). They performed rituals inside great kivas but collapsed, burned, and retired these large subterranean communal structures in the late 900s and used outdoor open plazas during the Classic (Creel and Anyon Reference Creel and Anyon2003). The Classic period is also defined by the presence of what archaeologists call Style III Classic Black-on-white pottery (Brody Reference Brody2004; Brody et al. Reference Brody, Scott and LeBlanc1983). Potters made black-on-white pottery before the Classic (Style I and II), but Style III has more intricate, fine-line, geometric designs and naturalistic images of humans, animals, and anthro- and zoomorphic figures, including turkeys.
Population estimates vary, but approximately 6,000 people lived in the entire Mimbres region, although several thousand fewer probably lived in the Mimbres Valley at its peak during the Classic (Blake et al. Reference Blake, LeBlanc and Minnis1986). At least a dozen pueblo villages with 100 or more rooms were built along the Mimbres River and its tributaries, but people also lived in smaller pueblos. Old Town, at the southern end of the valley, and Galaz, farther north, were economically and ritually significant places due to hundreds of years of occupation, great kiva precincts, and exotic objects such as copper bells, marine shell, and scarlet macaws (Anyon and LeBlanc Reference Anyon and LeBlanc1984; Creel Reference Creel, Powell-Martí and Gilman2006). Classic period Mimbres society was middle range and egalitarian, given that archaeologists have found little evidence for elites and centralized authority (Gilman Reference Gilman1990). However, beginning in the Late Pithouse period (550–1000) and continuing into the Classic, some individuals and families had social status and specialized leadership roles through access to prime agricultural land and esoteric ritual knowledge (Creel Reference Creel, Powell-Martí and Gilman2006; Roth and Baustian Reference Roth and Baustian2015; Shafer Reference Shafer, Powell-Martí and Gilman2006). Also, whereas people, pottery styles, and goods moved in and out of the valley during the Late Pithouse period, there was a relatively homogenized material culture during the Classic, with few nonlocal pottery types entering the valley, and people were more inward focused and isolated (Hegmon Reference Hegmon2002:339).
As was the case in the rest of the SW/NW during the 1000s and 1100s, maize was the primary food staple for people in the Mimbres Valley (Minnis Reference Minnis1985; see also Coltrain et al. Reference Coltrain, Janetski and Carlyle2007:Table 2). They also supplemented their diet with other domesticated crops and wild plants, and they hunted small and large game (Minnis Reference Minnis1985; Schollmeyer et al. Reference Schollmeyer, Diehl, Sandor, Roth, Gilman and Anyon2018). However, access to large game was unreliable during the Classic because artiodactyl populations had already been depleted by the Late Pithouse period (Cannon Reference Cannon2000; Nelson and Schollmeyer Reference Nelson and Schollmeyer2003). Finally, much of the Classic period experienced above-average rainfall, but a dry period from the 1120s to 1140s exacerbated environmental stress and population pressures (Grissino-Mayer et al. Reference Grissino-Mayer, Baisan and Swetnam1997; Minnis Reference Minnis1985; Nelson and Schollmeyer Reference Nelson and Schollmeyer2003). The end of the Classic period around 1130 is associated with the cessation of Style III pottery manufacture and the depopulation of much of the valley when people moved elsewhere (Nelson et al. Reference Nelson, Hegmon, Kulow and Schollmeyer2006).
Turkeys in the Mimbres Valley
Physical Evidence
Wild turkeys today live in the upland areas of the Gila Forest at the north end of the Mimbres Valley, given that their natural range and habitat are mountain forests 1,829–3,658 m (6,000–12,000 ft.) in elevation (Shaw and Mollohan Reference Shaw, Mollohan and Dickson1992). In Schollmeyer and MacDonald's (Reference Schollmeyer and MacDonald2020) synthesis of faunal remains from 105 sites across southwestern New Mexico, archaeologists have found at least 106 turkeys from 10 sites in the Mimbres Valley. However, we note that turkeys from Elk Ridge, Lake Roberts, Old Town, and Wheaton-Smith are not included in their study. The relative lack of turkey bones recovered from Mimbres Valley sites has puzzled archaeologists for nearly a century. For example, after excavating Swarts in the 1920s, Cosgrove and Cosgrove (Reference Cosgrove and Burton Cosgrove1932:5) noted that “with the exception of one bone fragment of the tarsometatarsus of a male bird there was an entire absence of turkey bones, either in the form of artifacts or as refuse. This is difficult to explain.” Because turkeys are uncommon in most Mimbres Valley avifaunal assemblages, few researchers have focused on turkeys in this region. However, more is known about turkeys from Elk Ridge, NAN Ranch, and Wind Mountain, and we provide summaries below.
Munro (Reference Munro and Smith2011:545) lists several archaeological signatures to infer that people managed turkeys. These include the presence of pens, dung deposits, eggshells, gizzard stones, fertilized eggs, young poults, turkeys with healed bone injuries, and an indication that the birds consumed maize. Many of these signatures are missing at most sites in the Mimbres Valley, but Morrison (Reference Morrison and Beckett2002) documented several of these signatures after studying 17 turkeys excavated from Elk Ridge's southern Classic period room block. In his preliminary analysis, Morrison (Reference Morrison and Beckett2002) estimated a 2:1 ratio of males (toms) to females (hens), and most were young adults between one and two years old. There were at least two adult birds of two years or older and one juvenile of three to five months. Three turkeys sustained broken and healed bone injuries, and eggshell fragments were found in room contexts. Also, archaeologists recovered partial pottery vessels on the room floors with the turkey remains. Partial pottery vessels in context with turkey remains have also been found outside the Mimbres Valley at Arroyo Hondo and Homol'ovi III (Lang and Harris Reference Lang and Harris1984:103; Senior and Pierce Reference Senior and Pierce1989:249). At Arroyo Hondo, Lang and Harris (Reference Lang and Harris1984:103) posit that people confined turkeys in these rooms and placed broken vessels on the floor to hold water.
The Elk Ridge turkey bones lack cut marks and burning, and archaeologists found articulated skeletons, suggesting that turkeys were not processed for consumption (Senior and Pierce Reference Senior and Pierce1989:251). Morrison (Reference Morrison and Beckett2002) also states that the birds were not ritually interred, because they were recovered from above the floor rather than below, where formal burials would be expected. However, recent excavations at the northern Classic period room block by Roth (Reference Roth2018) have yielded additional turkey remains, including birds buried in pits and roof fall. One such turkey was recovered from a small cobble-lined storage room that may have been a pen. Four additional articulated and partially articulated turkeys were also found on an extramural surface or possible ramada feature. No dung deposits or eggshells were recovered from these features.
Ten turkeys were identified at NAN Ranch. Eight were intentionally buried, including one that was buried with a bird effigy pendant and located adjacent to the burial of a child (Shaffer Reference Shaffer1991:Table 8.1). The remaining two turkeys were recovered from midden deposits. Shaffer (Reference Shaffer1991:127) offers that people at NAN Ranch, as at Elk Ridge, used turkeys for ritual rather than food. During excavations, Shafer (Reference Shafer2003:65, Figure 5.15) found a small walled feature that may have acted as a turkey pen, but archaeologists did not find bones, dung, or eggshells.
At Wind Mountain, located approximately 50 km west of NAN Ranch in the Gila region, turkeys were the most common avian species, with 178 bone fragments (Olsen and Olsen Reference Olsen, Olsen, Woosley and McIntyre1996). Six turkeys were intentionally buried, and one turkey femur had transverse knife cuts, indicating to Olsen and Olsen (Reference Olsen, Olsen, Woosley and McIntyre1996:397) that it had been butchered. Due to numerous immature bones, they argue that the turkeys were domesticated, but no poults, eggshells, pens, or dung deposits were recovered (Olsen and Olsen Reference Olsen, Olsen, Woosley and McIntyre1996:397, 400).
Iconographic Evidence
The Mimbres Valley is well known for the enigmatic black-on-white pottery made during the Classic period (Brody Reference Brody2004; Brody et al. Reference Brody, Scott and LeBlanc1983). Mimbres potters painted stylized designs of humans and animals sometimes engaged in various activities, and so researchers have used these images to understand aspects of Classic period Mimbres society (Hegmon et al. Reference Hegmon, Russell, Baller, Peeples and Striker2021; Hegmon and Trevathan Reference Hegmon and Trevathan1996; Munson Reference Munson2000). Therefore, no discussion of turkeys in the Mimbres Valley would be complete without briefly describing turkeys depicted on Mimbres Classic Style III Black-on-white pottery.
Classic period potters depicted anatomical features of several animals, and researchers have identified animals to the genus or species level, including some bird and fish species (Creel and McKusick Reference Creel and McKusick1994:512–515; Jett and Moyle Reference Jett and Moyle1986). Turkeys are identifiable on Mimbres painted pottery because potters included the turkey's snood, beard, wattle, or spur. Using various publications and the Mimbres Pottery Images Digital Database (MimPIDD; available online at https://core.tdar.org/collection/22070/mimbres-ceramic-database-with-search), we have identified 29 vessels with images of turkeys (Supplemental Table 1). We refer to each MimPIDD vessel using its Mimbres Archive (MA) number. We can distinguish turkeys from other birds based on the presence of the snood, beard, wattle, or spur, but we cannot discern if potters intended some birds to be turkeys but did not include these features. Also, although toms have longer or more pronounced snoods, beards, wattles, and spurs, some birds depicted on the vessels may be hens but lack these features. However, wild hens have snoods and wattles, albeit shorter and less pronounced, and some even have beards and spurs, although these are uncommon (Pelham and Dickson Reference Pelham, Dickson and Dickson1992).
Turkeys are the only naturalistic image on 59% of the vessels (n = 17, e.g., Figure 2A), but they share space on the remaining 41% (n = 12) with other animals and humans. For example, four vessels depict turkeys eating invertebrates (MA4001, MA4777, MA10317, and O'Bagy Davis Reference O'Bagy Davis1995:181). Also, five vessels have turkeys with humans in what appear to be narrative scenes. On MA5843 and MA9379 (Figure 3), men are depicted with turkeys, but potters did not include their anatomical sexual features on MA5651, MA5887, and in Fewkes (Reference Fewkes1923:Plate 11). However, they have objects and clothing most commonly associated with men (Munson Reference Munson2000), such as bows and arrows, given that they are hunting or capturing presumably wild turkeys. Some men wear turkey feathers (MA5651 and MA5843) and bird masks while hunting or stalking (MA5887). As shown in Figure 3 (MA9379), a man is wearing a feather in his hair and holding a maize stalk toward the turkey. He also has a staff or stick-like object at his feet.

Figure 3. Mimbres bowl (MA9379) depicting a man holding a maize stalk next to a turkey. Collection of the El Paso Museum of Archaeology, El Paso, Texas (# 95.005.002). (Vessel illustration by Caitlin Wichlacz. Vessel also illustrated in Fewkes [Reference Fewkes1924:Plate 42].)
Based on the presence of articulated turkey skeletons at some sites in the Mimbres Valley, people in this region used turkeys to some degree. Moreover, potters even depicted turkeys on their pottery vessels. Therefore, it is clear that these birds had some significance to Classic period Mimbres society. However, did Mimbres aviculturists have haplogroup H1 or H2 turkeys, and did the birds have C3, C4, or mixed diets?
Materials and Methods
Sampled Turkeys
The 31 turkey bones were recovered from Classic period subsurface contexts at Elk Ridge (n = 17), Lake Roberts (n = 3), Mattocks (n = 7), Old Town (n = 2), and Wheaton-Smith (n = 2). We chose these sites because they were professionally excavated, and the bones were more accessible for analysis than other collections. See Supplemental Text 1 for site summaries and Supplemental Table 2 for provenience information on the turkey samples.
Using McCaffery and colleagues (Reference McCaffery, Tykot, Gore and DeBoer2014:343) as a guide, turkey bones were selected based on preservation quality, size, and weight. To avoid sampling the same turkey twice, we chose the most common appendicular element and selected specimens from one side. We then repeated for the next most common element. However, it was sometimes difficult to use this strategy given that the five sites had limited availability of turkeys, or the bones had missing diagnostic features.
mtDNA and Stable Isotope Methods
See Supplemental Text 2 for the methods used for mtDNA extraction and sequencing (see also Supplemental Tables 3–5). The 31 turkey bones were analyzed at two ancient DNA laboratories. Twenty-four turkeys (Elk Ridge [n = 10], Lake Roberts [n = 3], Mattocks [n = 7], Old Town [n = 2], and Wheaton-Smith [n = 2]) were analyzed at Arizona State University (ASU), and seven Elk Ridge turkeys were analyzed at the University of Oxford. All turkey samples were extracted in dedicated ancient DNA cleanrooms using similar personal protective equipment to prevent contamination by modern DNA.
We used isotope values of bone collagen (δ13Cco, δ15Nco) to reconstruct dietary proteins of the 31 turkeys. See Supplemental Text 3 for the methods used. Bone collagen extraction and isotopic analyses were prepared at two laboratories. Twenty-four turkeys (Elk Ridge [n = 10], Lake Roberts [n = 3], Mattocks [n = 7], Old Town [n = 2], and Wheaton-Smith [n = 2]) were analyzed at the University of Florida, and seven Elk Ridge turkeys were analyzed at ASU. The results are expressed in parts per mil (‰) and delta (δ) notation. δ13C values are reported relative to the Vienna Pee Dee Belemnite (VPDB) fossil international standard, and δ15N values are reported relative to the atmospheric air (AIR) international standard.
Results
mtDNA
Previous mtDNA studies demonstrate that most turkeys from the northern Southwest belong to haplogroup H1, with fewer H2 turkeys (Kemp et al. Reference Kemp, Judd, Monroe, Eerkens, Hilldorfer, Cordray, Schad, Reams, Ortman and Kohler2017; Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010). Less is known about the maternal genetic lineage of turkeys from the southern Southwest or Northwest Mexico. However, two turkeys from the Gila Cliff Dwellings (TU1017 and 1019) in southwestern New Mexico and one (TU135) from the Calderón site in northern Chihuahua are haplogroup H2 (Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010:Table S3). Based on the extensive data from the northern Southwest and the scant data from farther south, it is difficult to hypothesize if the Mimbres turkey population will be primarily haplogroup H1 or H2. Therefore, this study contributes significantly to our understanding of turkey management as we increase the sample size of analyzed turkeys.
Of the 31 turkey samples we submitted for mtDNA analysis, 58% (n = 18) had sufficient mtDNA sequences to perform haplogroup calling. Unfortunately, 42% (n = 13) of the samples did not contain detectable mtDNA despite repeated extractions and polymerase chain reaction amplification (Elk Ridge [n = 2], Lake Roberts [n = 3], Mattocks [n = 6], and Old Town [n = 2]). Nevertheless, 56% (n = 10) of the successfully sequenced samples belong to the H1 haplogroup, and 44% (n = 8) are from the H2 haplogroup (Table 1). All 10 haplogroup H1 turkeys are from Elk Ridge, whereas the eight haplogroup H2 turkeys are from Elk Ridge (n = 5), Mattocks (n = 1), and Wheaton-Smith (n = 2). Based on this study, Mimbres aviculturists used their haplogroup H2 population almost as much as their haplogroup H1 population, unlike in the northern Southwest.
Table 1. Summary Table of Genetic and Isotope Results.

We observed 11 different mitochondrial haplotypes from the 18 turkeys (see Supplemental Figures 1, 2, and 3). Haplotypes from haplogroup H1 include aHap1e, aHap1f, and the most common, aHap1. Previous mtDNA studies did not identify haplotypes aHap1e and aHap1f. The turkeys belonging to haplogroup H2 seem to be more diverse than what researchers have found elsewhere, because five new haplotypes are in the Mimbres sample (aHap2f, aHap2g, aHap2w, aHap2y, and aHap2z). The presence of these five haplotypes can be explained in several ways, including sampling biases, lack of complete mtDNA genome coverage, or a more historically based reason such as an influx of turkeys that had yet to be completely domesticated in the Mimbres Valley. We also identified two previously recognized haplotypes in modern Merriam's turkeys (aHap2h and aHap2x), both from Elk Ridge. Furthermore, we identified one haplotype aHap2c turkey from Mattocks. Speller and colleagues (Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010:Table S3) identified this haplotype at McPhee Village in Colorado (TU21) dating to 820–980, the Gila Cliff Dwellings (TU1019) dating to 1270–1290, and Point of Pines in Arizona (TU1101) dating to 1200–1400. Lipe and colleagues (Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:Table 1) also identified one haplotype aHap2c turkey from the Croom site in Utah (GG-145.41.1) dating to 620–700. This information suggests that haplotype aHap2c turkeys were present across an extensive geographic range for several centuries.
δ13C and δ15N Isotopes
Previous studies of turkey diet in the SW/NW demonstrate that most consumed maize, a C4 plant, but some consumed nonmaize C3 plants or even had a mixed diet (Jones et al. Reference Jones, Conrad, Newsome, Kemp and Kocer2016; Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016; Manin et al. Reference Manin, Corona-M, Alexander, Craig, Thornton, Yang, Richards and Speller2018; McCaffery et al. Reference McCaffery, Tykot, Gore and DeBoer2014, Reference McCaffery, Miller and Tykot2021; Rawlings and Driver Reference Rawlings and Driver2010; Webster and Katzenberg Reference Webster, Anne Katzenberg and Kemrer2008). Therefore, we should expect similar results for the Mimbres turkeys. However, based on Mimbres iconography, turkeys likely consumed invertebrates and maize. Of the 31 turkey samples we submitted for stable isotope analyses, we obtained results for 94% (n = 29) of the samples (Table 1). However, one turkey from Mattocks (MT07) and one from Old Town (OT02) had carbon to nitrogen (C:N) ratios outside the acceptable range of reasonably well-preserved collagen: 2.9–3.6 (DeNiro Reference DeNiro1985). These two samples are not included in Figures 4 and 5, but these data can be found in Supplemental Table 6.

Figure 4. Bone collagen δ13C and δ15N isotope values from Elk Ridge, Lake Roberts, Mattocks, Old Town, and Wheaton-Smith turkeys.
The 29 Mimbres turkeys exhibit δ13C values ranging from −16.9‰ to −6.9‰ (−9.4‰ avg.), and the δ15N values range from 3.6‰ to 10.2‰ (7.8‰ avg.). Rawlings and Driver (Reference Rawlings and Driver2010:2435) note, “Animals with diets that are composed primarily of C3 plants generally express average δ13C values between −22‰ to −19‰, while those consuming primarily C4 plants have values that range between −10‰ to −6‰.” Diets consisting of a mixed feeding ecology have values between −18‰ and −11‰ (Manin et al. Reference Manin, Corona-M, Alexander, Craig, Thornton, Yang, Richards and Speller2018; Tykot Reference Tykot, Staller, Tykot and Benz2006). As for δ15N values, herbivores have values averaging between 6‰ and 9‰, carnivores have higher values, and omnivores have intermediate values (McCaffery et al. Reference McCaffery, Tykot, Gore and DeBoer2014; Rawlings and Driver Reference Rawlings and Driver2010; Webster and Katzenberg Reference Katzenberg, Anne Katzenberg and Sanders2008). McCaffery and colleagues (Reference McCaffery, Miller and Tykot2021:Table 3) also provide estimated values. They suggest that turkeys with C4 diets have δ13C values of −10.20‰ ± 0.62 (SD) and δ15N values of 6.17‰ ± 2.46 (SD). On the other hand, turkeys with C3 diets have δ13C values of −24.10‰ ± 2.78 (SD) and δ15N values of 1.38‰ ± 3.15 (SD). They also provide values for turkey diet consisting of C4 invertebrates (δ13C −10.70‰ ± 0.62 [SD], δ15N 8.05‰ ± 2.46 [SD]) and C3 invertebrates (δ13C −24.60‰ ± 2.78 [SD], δ15N 3.26‰ ± 3.15 [SD]).
Using these values as a guide, we determined that 79% (n = 23) of our sample exhibited δ13C values indicating that the turkeys consumed C4 plants, 14% (n = 4) had δ13C values indicating that they consumed C3 plants, and the remaining 7% (n = 2) consumed a mixed diet. The average δ13C value for C4 consumers is −8‰ (range −9.9‰ to −6.9‰), the average δ13C value for C3 consumers is −16.8‰ (range −16.9‰ to −15.3‰), and the average δ13C value for turkeys with mixed diets is −11.3‰ (range −11.6‰ to −11‰). The two mixed-diet turkeys (OT01 and WS02) have δ13C values closer to C4 consumers than C3 consumers. Based on our comparison of the 29 Mimbres turkeys to 269 other archaeological turkeys from the SW/NW (n = 232) and southeastern United States (n = 37), 13 modern turkeys, and 215 ancient humans from the SW/NW in Figure 5, the C4 Mimbres turkeys align with most of the other SW/NW ancient turkeys and humans. The C3 Mimbres turkeys have values similar to the other C3-consuming ancient SW/NW, southeastern, and modern turkeys.

Figure 5. Bone collagen δ13C and δ15N isotope values from the 29 Mimbres turkeys reported in this article compared to 232 archaeological turkeys from the SW/NW (Conrad et al. Reference Conrad, Jones, Newsome and Schwartz2016:Appendix A; Jones et al. Reference Jones, Conrad, Newsome, Kemp and Kocer2016:Table 3; Kellner et al. Reference Kellner, Schoeninger, Spielmann, Moore and Morgan2010:Table 8.3; Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:Table 1; Manin et al. Reference Manin, Corona-M, Alexander, Craig, Thornton, Yang, Richards and Speller2018:Table S1; McCaffery et al. Reference McCaffery, Tykot, Gore and DeBoer2014:Table 2; McCaffery et al. Reference McCaffery, Miller and Tykot2021:Table 4; Rawlings and Driver Reference Rawlings and Driver2010:Tables 5, 6), 37 archaeological turkeys from the southeastern United States (Thornton et al. Reference Thornton, Peres, Chase, Kemp, Frome, Manin, Basnett, McGrath, Speller and Reitz2021:Table S1), 13 modern turkeys (Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:Table 2), and 215 ancient humans from the SW/NW (Coltrain and Janetski Reference Coltrain and Janetski2013:Tables 1, 2, 3; Coltrain et al. Reference Coltrain, Janetski and Carlyle2007:Table 1; Martin Reference Martin1999:Table 2).
Combining mtDNA and Isotope Data
When possible, it is helpful to combine genetic and dietary data on individual turkeys to best understand management practices (Jones et al. Reference Jones, Conrad, Newsome, Kemp and Kocer2016; Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016; Manin et al. Reference Manin, Corona-M, Alexander, Craig, Thornton, Yang, Richards and Speller2018; Thornton et al. Reference Thornton, Peres, Chase, Kemp, Frome, Manin, Basnett, McGrath, Speller and Reitz2021). We obtained genetic and dietary data for 18 turkeys from Elk Ridge (n = 15), Mattocks (n = 1), and Wheaton-Smith (n = 2). Ten haplogroup H1 turkeys consumed a C4 diet, one haplogroup H2 turkey consumed a C3 diet, six haplogroup H2 turkeys consumed a C4 diet, and one haplogroup H2 turkey had a mixed diet.
As shown in Figure 6, the δ13C and δ15N values of the 10 haplogroup H1 Mimbres turkeys are more tightly clustered than the eight haplogroup H2 turkeys. To further investigate this, we used a measurement of central tendency and dispersion to compare the two haplogroups. We used nonparametric measures of the median and interquartile ranges due to the small sample sizes and because the data are not normally distributed. As shown in the box and whisker plots in Figure 7, the interquartile ranges of δ13C and δ15N are much broader for the haplogroup H2 turkeys than for haplogroup H1 turkeys. This confirms that the stable isotope values of the haplogroup H2 turkeys are more variable and widely dispersed. To determine if this pattern occurs elsewhere, we compared the isotope values of the 10 Mimbres and 28 SW/NW haplogroup H1 turkeys to the 8 Mimbres and 11 SW/NW haplogroup H2 turkeys (Figure 8). Except for the four haplogroup H1 turkeys reported by Jones and colleagues (Reference Jones, Conrad, Newsome, Kemp and Kocer2016) that consumed C3 diets—shown in the bottom-left corner of Figure 8—the values of the haplogroup H1 turkeys are more tightly clustered together than the values of the haplogroup H2 turkeys. Therefore, whereas Mimbres and Ancestral Pueblo aviculturists provided maize to haplogroup H1 and H2 turkeys, the haplogroup H2 birds had somewhat more varied diets.

Figure 6. Bone collagen δ13C and δ15N isotope values from the haplogroup H1 and H2 Mimbres turkeys.

Figure 7. Box and whisker plots comparing the median and interquartile ranges of δ13C and δ15N values for haplogroups H1 and H2. The shaded boxes display the interquartile range where 50% of the δ13C and δ15N values plot in relation to the median (bold line). The circles marked with a turkey sample number (ER04 and ER08) denote statistical outliers.

Figure 8. Bone collagen δ13C and δ15N isotope values from the 10 Mimbres and 28 SW/NW haplogroup H1 turkeys (Jones et al. Reference Jones, Conrad, Newsome, Kemp and Kocer2016:Table 3; Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:Table 1) compared to the eight Mimbres and 11 SW/NW haplogroup H2 turkeys (Jones et al. Reference Jones, Conrad, Newsome, Kemp and Kocer2016:Table 3; Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:Table 1).
Discussion
This study aims to understand turkey management practices in the Mimbres Valley during the Classic period using mtDNA and stable carbon and nitrogen isotope analyses. People in the Mimbres Valley had haplogroup H1 and H2 turkeys, and the birds primarily consumed a C4 diet, but some had a nonmaize C3 diet, and even fewer had a mixed diet. Elk Ridge is the only site in our sample with evidence of both haplogroups. By integrating the genetic and dietary data, we see that Mimbres aviculturists treated haplogroup H1 and H2 birds alike, which means that most were fed C4 diets, which is similar to the way Ancestral Pueblo groups managed their turkeys. Data also show that haplogroup H2 Mimbres turkeys had more varied diets than haplogroup H1 turkeys, a pattern also seen in the northern Southwest.
We are uniquely positioned to examine key issues in Mimbres archaeology because this study is the first to investigate the maternal genetic lineage and diet of turkeys from Classic period sites. Below, we address how the human-turkey relationship began in this region, the evidence for turkey pens and reducing turkey mobility, and the socioecological factors related to turkey management—notably, the challenges associated with feeding maize to turkeys during times of environmental stress. Answering these questions provides new insights into Mimbres society and how turkeys fit within Classic period subsistence, economic production, sociopolitical organization, and religious and ritual practice.
The Human-Turkey Relationship
The human-turkey relationship in the Mimbres Valley likely began because of an increasing population that aggregated into pueblo villages along the Mimbres River. People cleared forests to make additional space for agricultural fields, and trees were needed for construction beams and fuel for fires. Clearing forests destroyed wild turkey habitat, and people may have inadvertently attracted turkeys closer to their maize farming villages. Turkeys exhibit several favorable traits for domestication and management of genetically wild birds, including a social structure based on a dominance hierarchy, a tolerance for penning, flocking behavior, and a generalist diet (Zeder Reference Zeder2012a:Figure 1). Turkeys are also precocial birds, which means that they are well developed, are down covered, and can stand and walk right after hatching (Healy Reference Healy and Dickson1992). Newly hatched poults imprint on the first animal providing parental care, and the birds respond to that parent—even humans—as if it were a turkey. They are relatively social birds, and like dogs, turkeys are commensal domesticates because they take advantage of the food and shelter people provide (Zeder Reference Zeder, Gepts, Famula, Bettinger, Brush, Damania, McGuire and Qualset2012b:240–242). Wild turkeys are initially timid, but they would have benefited from the food resources around Mimbres farming villages once they became familiar with the sights and sounds of being close to humans.
The Mimbres genetic and dietary data corroborate Lipe and colleagues’ (Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:108–109) position that viewing haplogroup H1 and H2 turkeys as “domestic” versus “wild” is an oversimplification. Despite their genetic differences, Ancestral Pueblo and Mimbres aviculturists provided maize to haplogroup H1 and H2 turkeys. Nevertheless, there is a clear difference in the frequency of haplogroup H1 and H2 turkeys in the northern Southwest with far more from the H1 lineage than the H2 lineage (Kemp et al. Reference Kemp, Judd, Monroe, Eerkens, Hilldorfer, Cordray, Schad, Reams, Ortman and Kohler2017; Speller et al. Reference Speller, Kemp, Wyatt, Monroe, Lipe, Arndt and Yang2010). However, based on this Mimbres study, people in the valley relied almost as much on their haplogroup H2 population as their haplogroup H1 population. Furthermore, unlike other areas of the SW/NW, people in the Mimbres Valley did not have a history of large-scale prehispanic turkey use. How then did people in this region acquire turkeys?
Mimbres aviculturists likely captured haplogroup H2 birds in the wild to increase their turkey population, which is similar to what residents of historic Taos Pueblo in northern New Mexico who confined wild-caught turkeys at the pueblo did (Schorger Reference Schorger1961:140). Also, as Lipe and colleagues (Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:109) mention, the haplogroup H2 birds were not themselves wild, given that humans managed them and provided maize, but they were likely descendants of wild turkeys that were integrated into flocks and that gave rise to future generations of haplogroup H2 birds. As for haplogroup H1 turkeys, Elk Ridge is currently the only Classic period site in the Mimbres Valley with evidence of this lineage. People at Elk Ridge could have acquired haplogroup H1 eggs or poults from northern groups. Future studies could use radiogenic strontium isotopes to investigate if people acquired turkeys from outside the valley (Grimstead et al. Reference Grimstead, Reynolds, Hudson, Akins and Betancourt2016; Schwartz et al. Reference Schwartz, Somerville, Nelson and Knudson2021).
Our study validates Morrison's (Reference Morrison and Beckett2002) position that Elk Ridge was a localized experiment in turkey management because people there likely had the knowledge, technology, and interest to raise and keep these birds. Perhaps people at Elk Ridge learned aviculture from more northern groups who had centuries of experience managing turkeys. Other people in the Mimbres Valley may have visited Elk Ridge or traded with them to acquire turkey feather blankets, robes, or the feathers themselves. Elk Ridge also has evidence of pottery production (Creel and Speakman Reference Creel, Speakman, Roth, Gilman and Anyon2018). Consequently, people there played an essential role in the movement of goods within the larger Mimbres Valley. People who managed macaws in Mimbres society likely had higher social status (Creel and McKusick Reference Creel and McKusick1994; Munson Reference Munson2000), so it is probable that those who managed Elk Ridge's turkeys also had specialized leadership roles in the village, given that it took special knowledge to manage birds. People at Elk Ridge managed turkeys, but is there evidence they used pens or other means of reducing turkey mobility?
Mimbres Valley Turkey Pens
People corralled turkeys in pens or kept them in abandoned rooms to restrict the birds’ mobility, control their breeding, and safely keep them to acquire important resources such as feathers and meat (Conrad Reference Conrad2022). Pens are uncommon in the archaeological record because people built them using materials that do not often preserve, such as wooden poles, branches, and adobe. Some of the best examples of pens are from Pindi Pueblo, Arroyo Hondo, and Paquimé, and people at these sites had up to several hundred turkeys (Di Peso et al. Reference Di Peso, Rinaldo and Fenner1974; Lang and Harris Reference Lang and Harris1984; Stubbs and Stallings Reference Stubbs and Stallings1953).
Penned turkeys were regularly fed maize and protected from predators, but they arguably lived a more miserable life than their wild cousins. Penning for long intervals was damaging to the birds’ health because they were confined in small spaces, stressed, and surrounded by excrement. There were also human consequences for keeping large numbers of birds penned within the pueblo village. As with macaws, the confinement of turkeys could have spread zoonotic diseases such as psittacosis throughout the village (Schwartz et al. Reference Schwartz, Taylor, Hegmon, Schwartz, Plog and Gilman2022:311–312). Mimbres aviculturists could have contracted psittacosis by inhaling dust particles contaminated with the birds’ fecal matter. If untreated, humans can experience high fever, diarrhea, respiratory infection, and death.
Archaeologists have not found definitive evidence of formal turkey pens in the Mimbres Valley. However, aviculturists at Elk Ridge likely kept the birds in unused rooms, and there are small rooms at NAN Ranch and Old Town that may have been used to confine turkeys (Creel et al. Reference Creel, Cannon, Broughton, Francis and Ludeman2015:6; Shafer Reference Shafer2003:65). We recommend that archaeologists use Conrad (Reference Conrad2022) as a guide during future excavations at Classic period Mimbres pueblo sites to help look for turkey pens. Nevertheless, people in the Mimbres Valley may not have penned their turkey population because they had relatively few birds compared to other sites, such as Paquimé. However, below, we provide evidence to suggest that people at Elk Ridge limited turkey mobility. People at NAN Ranch and Old Town may also have limited mobility, but more work is needed to understand turkey management at those sites.
The first line of evidence comes from the potential watering bowls on the floors of Elk Ridge rooms with turkeys. Rather than constructing separate formal pens, people at Elk Ridge may have kept turkeys in unused rooms and placed broken pottery vessels on the floor so they could drink. If turkeys were kept in those rooms, we should expect thick layers of yellowish soil. However, excavations did not uncover dung deposits at Elk Ridge, and dung deposits have not been found at other Mimbres sites (Creel et al. Reference Creel, Cannon, Broughton, Francis and Ludeman2015:6). It is possible, however, that people could have cleaned the room(s) of dung, as Conrad (Reference Conrad2022:647) proposes, or they could have tethered turkeys in different locations entirely, such as extramural areas outside of the pueblo.
The second line of evidence for limiting turkey mobility is from the presence of healed bone injuries on three male turkeys at Elk Ridge. According to Morrison (Reference Morrison and Beckett2002), one suffered a broken left tibiotarsus that had healed prior to the bird's death, another had a severe break on the right humerus, and the third had a healed pathology on the left tibiotarsus. These birds most likely would not have survived in the wild, requiring human care and management. It is possible that these birds received their injuries from being tethered because tethering was a common management practice (Conrad Reference Conrad2022; Fothergill Reference Fothergill2016). Mimbres aviculturists may have tethered or leashed the birds to heavy objects such as rocks or trees. Besides the trauma indicated on bones, archaeological evidence for tethering is rare because the cord tied to the bird does not preserve in most conditions. There are rare instances of desiccated turkeys with the cord still attached, but not from the Mimbres Valley (Lipe et al. Reference Lipe, Kyle Bocinsky, Chisholm, Lyle, Dove, Matson, Jarvis, Judd and Kemp2016:106).
Socioecological Interactions
How people in the Mimbres Valley interacted with their environment is one of the more prominent themes in Mimbres archaeology. For example, much is known about Classic period landscape modification, hunting, farming, and gathering strategies, and how these actions impacted resource availability (Minnis Reference Minnis1985; Nelson and Schollmeyer Reference Nelson and Schollmeyer2003; Nelson et al. Reference Nelson, Hegmon, Kulow and Schollmeyer2006; Schollmeyer Reference Schollmeyer2011; Schollmeyer and Driver Reference Schollmeyer and Driver2013). However, how did turkey management practices impact socioecological aspects of Classic period Mimbres society related to ancient food webs, farming practices, and potential food insecurities?
During the last few decades of the Classic period, farmers had difficulty growing crops because there was a dry period, the soils were degraded, and floodplain vegetation was depleted due to centuries of intense land use (Minnis Reference Minnis1985; Sandor et al. Reference Sandor, Hawley, Schiowitz, Gersper, Mack, Witcher and Lueth2008). Nevertheless, Mimbres farmers could have still produced enough crops to survive, but they adapted by using less productive fields or travel to more distant fields (Schollmeyer Reference Schollmeyer2011). Also, people lacked reliable protein from large mammals because artiodactyl populations were already reduced due to landscape modification and overhunting (Cannon Reference Cannon2000; Nelson and Schollmeyer Reference Nelson and Schollmeyer2003). Consequently, people during the Classic may have needed to find new animal protein sources to avoid potential food insecurities.
Animals are costly to keep because humans must provide food, shelter, and care, or else the animal will become sick, die, and not produce the valuable resources humans want. As in much of the SW/NW, people in the Mimbres Valley fed turkeys maize. Keeping turkeys was costly for maize agriculturists because people and their turkeys relied on the same crop to survive (Bocinsky and Kohler Reference Bocinsky, Kohler, Thomas Foster, Paciulli and Goldstein2015). If people in the Mimbres Valley went through times of environmental stress and had food shortages, they may have needed to make decisions about their turkey population. For example, they could have eaten their turkeys, similar to those in the Mesa Verde region in southwestern Colorado who began eating turkeys once their artiodactyl populations decreased (Badenhorst and Driver Reference Badenhorst and Driver2009; Ellyson et al. Reference Ellyson, Nagaoka and Wolverton2019). Wild toms can weigh approximately 7.5 kg (16.5 lb.), and mature hens can weigh 4.5 kg (10 lb.; Schorger Reference Schorger1966:88, Table 9). Consequently, eating turkeys would have offered more protein and fat while at the same time reducing the number of birds so as to conserve maize for human consumption. However, unlike those in the Mesa Verde region, Mimbres groups chose not to eat their turkeys even though they had unreliable access to artiodactyls, and maize farming may have been more difficult.
Why did Mimbres groups keep turkeys if not for a readily available protein source? It is most likely that people kept turkeys alive because they wanted the feathers. Feathers from several bird species were ritually important because they were added to prayer sticks that were deposited as gifts to ancestors to bring rain (Tyler Reference Tyler1979:93). In addition, turkey feather blankets are warmer than rabbit-fur blankets (Blinman Reference Blinman2001). Because turkey feathers were necessary for ritual and utilitarian purposes, it was more important for Mimbres society to keep live turkeys for their feathers rather than for food consumption. Although there is iconographic evidence of men hunting or capturing presumably wild turkeys—likely for their feathers rather than meat—people could have collected their feathers when they molted naturally or plucked feathers from live birds (Fothergill Reference Fothergill2016:564).
Conclusions
More than ever, archaeologists are studying how past peoples throughout North America used and managed turkeys. In this article, we used mtDNA and stable carbon and nitrogen isotope analyses to investigate turkey management practices in the Mimbres Valley from AD 1000 to 1130. Using these data, we provided new information about how turkeys fit within Classic period subsistence, economic production, sociopolitical organization, and religious and ritual practice. Like Ancestral Pueblo groups in the northern Southwest, Mimbres aviculturists managed haplogroup H1 and H2 turkeys and fed these birds primarily C4 maize diets. However, unlike Ancestral Pueblo groups, Mimbres aviculturists managed more haplogroup H2 turkeys and did not eat the birds (Morrison Reference Morrison and Beckett2002; Shaffer Reference Shaffer1991). Instead, feather production outweighed any gains from their use as food, even when turkey protein would have helped reduce food insecurity during parts of the Classic period.
This study has broad relevance to places where people managed wild, tame, and domestic animals, and we offered new insights into how prehispanic, small-scale, middle-range agricultural societies in the SW/NW managed turkeys for ritual and utilitarian purposes. This study also provides essential baseline information on turkey management for the Mimbres Valley, which can be used to compare the human-turkey relationship throughout North America. However, additional research on the maternal genetic lineage and diet is needed on turkeys from other sites and periods in the southern Southwest and Northwest Mexico to confirm if turkey management practices change over time and across space. Finally, the next decade will undoubtedly generate further insights on turkey management, with improved analytical methods and innovative research questions in collaboration with archaeologists, Native American communities, geneticists, and wildlife biologists.
Acknowledgments
We thank the following individuals and institutions for facilitating access to turkey samples: Karen Gust Schollmeyer, Michael Cannon, Steven LeBlanc, Julia Clifton and Diana Sherman (Center for New Mexico Archaeology / Museum of Indian Arts and Culture), Robin Cordero (formerly of Office of Contract Archeology / University of New Mexico), Jack Young (New Mexico Department of Game and Fish), Thomas Holcomb (Bureau of Land Management, Las Cruces office, retired), and Barbara Roth (University of Nevada, Las Vegas). We also thank Kyle Bocinsky, Katelyn Bishop, Patricia Gilman, and the anonymous reviewers who provided comments on earlier drafts. We appreciate Caitlin Wichlacz for illustrating Figures 2 and 3, and Myles Miller for providing statistical analyses and Figure 7.
Funding Statement
We thank the following organizations who funded this project: The Rust Family Foundation (RFF-2019-106, to Dolan and Laumbach and administered by Dr. Deborah Dennis, Human Systems Research Inc.), the New Mexico Archaeological Council (to Dolan), the Center for Evolution and Medicine at Arizona State University (to Ozga), and a European Research Council Grant to conduct a portion of the genomic analyses (MSCA-IF-2016-748679 TURKEY, to Manin).
Data Availability Statement
The data supporting this research are included in the electronic supplemental material. The turkey genetic sequences have been deposited to GenBank (http://www.ncbi.nlm.nih.gov/genbank/) under accession numbers OP056179-OP056196.
Supplemental Material
For supplemental material accompanying this article, visit https://doi.org/10.1017/aaq.2022.81.
Supplemental Figure 1. Coverage distribution on the 15567-16004 segment from the HTS data.
Supplemental Figure 2. Median-joining network displaying the relationships between the newly obtained sequences and previously published archaeological and modern sequences obtained from GenBank.
Supplemental Figure 3. Median-joining network displaying the relationships between the newly obtained sequences and previously published archaeological and modern sequences obtained from GenBank.
Supplemental Table 1. Turkey Images on Mimbres Pottery.
Supplemental Table 2. Turkey Provenience Information.
Supplemental Table 3. mtDNA Sample List.
Supplemental Table 4. Diagnostic Mutations Observed in the Sequences and Haplogroup Calling from the HTS and Sanger Data.
Supplemental Table 5. List of Samples Used in the Haplotype Calling.
Supplemental Table 6. Isotopic Results from This Study.
Supplemental Text 1. Archaeological Site Summaries.
Supplemental Text 2. mtDNA Analysis and Laboratory Methods.
Supplemental Text 3. Stable Carbon and Nitrogen Isotope Analysis and Laboratory Methods.
Competing Interests
The authors declare none.











