Chapter 1 Toward a scientific-realistic theory on the origin of human behavior
For almost half a century, anthropologists have debated to what extent their discipline can be considered a science, and if so, whether it should be viewed as a social science with different methods and heuristics from natural sciences (Binford, Reference Binford1962, 1968; Aberle, Reference Aberle, Binford and Binford1968; Clarke, Reference Clarke1968, 1972; Watson et al., Reference Watson, LeBlanc and Redman1971; Schiffer, Reference Schiffer1975; Gould, Reference Gould1978, 1980; Zeitlin, Reference Zeitlin1990; Wylie, Reference Wylie2002; Turner, Reference Turner2007). In the current state of affairs, and despite the firm belief that some anthropologists (mainly processualists) have that our discipline is a scientific endeavor, there are reasons to think that even processualists have not gone far enough to render their approach to anthropology fully scientific. The belief in systemics, the use of statistics, and the contrasting of null hypotheses are not enough to qualify as a science. A scientific undertaking requires a well-defined theory, articulated around a set of well-contrasted hypotheses in which every premise and assumption are clearly defined (Bunge, Reference Bunge1998a). Theories abound in archaeology; as a matter of fact it would not be an exaggeration to claim that there are as many theoreticians as there are field researchers these days (note that these terms should be complementary, not mutually exclusive). Most of these theories could be better served, however, by being described as theoretical positioning or nonscientific theories rather than scientific theories per se, because most of them lack well-articulated contrasting sets of hypotheses. One could subscribe to Marxist, historical-cultural, cultural-materialist, functionalist, or any other theoretical school; however, the main axioms of these schools’ theories remain untested, tested but not supported by evidence, or plainly not testable (Popper, Reference Popper1957, 1965, 1972; Bunge, Reference Bunge1998a, 1998b; Psillos, Reference Psillos1999).
Thus, anthropologists are often perceived by philosophers of science as researchers who are
[d]iffident regarding theory…making hypotheses, which they often dignify with the name of theories.…Actually, aside from vague “grand theories” such as evolutionism, diffusionism, functionalism, conflict theory and cultural materialism – all of them largely programmatic hypotheses that have inspired fruitful research projects – anthropology contains hardly any theories, that is, hypothetico-deductive systems.… The typical research project in anthropology is a fact-finding mission. (Bunge, Reference Bunge1998b, 47)
Some processualists (see Binford, Reference Binford2001) have strived to emphasize that programmatic questions produce hypotheses that must be tested against the empirical evidence of the archaeological record and interpreted under the analogical dynamics of referential frameworks. Theory thus should be of utmost relevance. Partly to accomplish this purpose to a certain degree, the middle-range theory was borrowed from Merton's (1967) work in sociology and articulated into several uniformitarian forms: ethnographic analogy, ethnoarchaeology, experimental archaeology, and neo-taphonomy, among others (Atici, Reference Atici2006). The systemic concept of human culture itself to which this methodology was applied (New Archaeology) was never scientifically tested in a successful way, however, but was inferred from patterns of relatedness among selected variables for which the available information is heterogeneous (Binford, Reference Binford2001). This differs from assumptions derived from systemic relations among selected behavioral variables phrased in the form of testable hypotheses, with their corresponding falsifying premises. This shortcoming should be emphasized, despite the acknowledgment that a systemic approach to human behavior is the only way to understand patterns and regularities in it (Bunge, Reference Bunge and Mahner1982, 1998b).
Although traditional critics and even some supporters of the processual approach emphasize its logical positivist foundation (Deetz, Reference Deetz1970; Flannery, Reference Flannery and Redman1973; Read and LeBlanc, Reference Read and LeBlanc1978), the rejection of the metaphysical components of theories, the emphasis on hypotheses not necessarily articulated over theories, and the necessity of justifying all of the assertions of interpretations empirically, as defended in the logical positivism philosophy, conflict with the emphasis that processualists in their early years placed on theories with all their metaphysical components (although one can argue that most of them were not necessarily scientifically articulated). The New Archaeology thus “diverged sharply from the deductivist models of explanation and confirmation associated with such latter-day exponents of logical positivism/empiricism as Hempel” (Wylie, Reference Wylie2002, p. 81). Furthermore, the conceptual schemes and procedures of processualists are better understood within “scientific realism, a theory of science that…offers a much more congenial framework for the New Archaeology than does Hempelian positivism” (Wylie, Reference Wylie2002, p. 24). I concur with Wylie (Reference Wylie2002) that processual anthropology better fits the epistemic criteria of scientific realism than those of positivism but argue that it is still far from adopting a scientific realistic framework for its praxeology.
Although much closer to a scientific approach than other theoretical approaches, the classical processualist approach, as is discussed later in this chapter, also lacks complete epistemological justification and does not quite fit into what is currently understood as a scientific theoretical body of knowledge, as is defined in scientific realism, which is the pragmatically dominant philosophical epistemic school in natural sciences (Putnam Reference Putnam, Pearce and Maynard1972, 1975; Bunge, Reference Bunge and Mahner1973, 1982, 1998b, Reference Bunge2006; Byerly and Lazara, Reference Byerly and Lazara1973; Toumela, Reference Toumela1973; Boyd, Reference Boyd1983; Miller, Reference Miller1987; Niiniluoto, Reference Niiniluoto1987, Reference Niiniluoto2002; Lipton Reference Lipton1993; Aronson et al., Reference Aronson, Harre and Cornell Way1995; Bhaskar, Reference Bhaskar1998, Reference Bhaskar2007; Psillos, Reference Psillos1999).
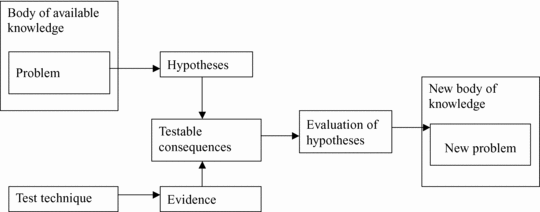
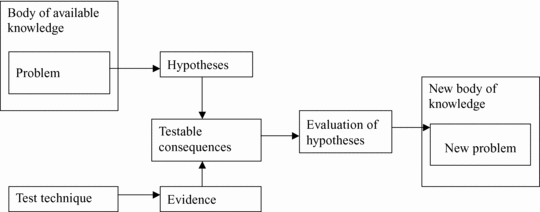
Scientific realism requires logic reasoning that is organized systematically. Scientific systematization implies the use of concepts within hypothetical constructs (containing inferable entities or properties), which should avoid vagueness and be properly defined if they are to be meaningful. These concepts should form theorems deduced from prior axioms or founder hypotheses. The systemic articulation of theorems or hypotheses containing theoretical well-defined concepts creates factual theories (Bunge, Reference Bunge1998a; Niiniluoto, Reference Niiniluoto2002). These systemic articulations (theories) constitute the body of knowledge within which problems at the root of research arise. These problems lead to new hypotheses with testable consequences that after testing can be evaluated, providing corroboration or rejection and a subsequent increased body of knowledge. This is what is epistemologically called the method of successive approximation (Bunge, Reference Bunge1998a), truthlikeness (Niiniluoto, Reference Niiniluoto1987), or lineal (evolutionary) progression of knowledge (Popper, Reference Popper1972). Inherent in this widespread conception of “theory” is the interpretation of theories as approximate images of reality containing webs of laws or patterns. This renders theories as modeled patterns of reality. Because the goal of a theory is not to provide an answer to exceptions and oddities, theories should be interpretive patterns explaining laws, regularities, and general processes of reality (Niiniluoto, Reference Niiniluoto1987; Psillos, Reference Psillos1999). Exceptions to these laws or patterns must be explained with auxiliary hypotheses. These hypothetic-deductive systems are the only ones that can generate enough background knowledge to provide new problems for research.
Such hypothetic-deductive systems are poorly defined in anthropology. In the case of anthropological research, theories are weak because they lack clear definitions with which to start. As Bunge (Reference Bunge1998b) acknowledges, the very first problem anthropologists face is the definition of their object of study: What is man? What is defined by human? Without clear definitions, there are no valid theoretical concepts, and therefore no theories. Regarding the latter question, an answer for a definition of human behavior from an evolutionary perspective was provided by Isaac (Reference Isaac1978) when he identified as human all the novel features that humans do not share with the rest of primates. This exemplified a widespread concept of interpretation in anthropology, which is using analogic reasoning following the uniformitarian principle of actualism (Binford, Reference Binford1962, 1978, 1981, 2001). What was missing from Isaac's diagnosis, however, was a way of applying those features to the archaeological record by means of a well-articulated scientific theory. A similar criticism could be applied to models of modern human behavior (reviewed in Henshilwood and Marean, Reference Henshilwood and Marean2003), which are based on analogically derived lists of ethnological and archaeological traits.
My modest goal here is to provide an example of an articulated theory for the origin of human behavior that could be used as the contrasting body of knowledge against which research problems can be created and potentially solved through palaeoanthropological research. Thus, a scientific approach to this issue could be more successfully defended than it has been so far and would encounter less epistemological criticism by philosophers of science subscribing to the realistic school of thought. To carry out this task, I intend to use formal logic and the principles of theory building as described by scientific realists (Niiniluoto, Reference Niiniluoto1987; Aronson et al., Reference Aronson, Harre and Cornell Way1995; Psillos, Reference Psillos1999), with special emphasis in the guidelines provided by a variant of scientific realism: hylorealism (Bunge, Reference Bunge1998a, 2006). I do not intend here to champion this approach over any other theoretical approach; my argument is that it is more scientifically sound than previous science-related (processual) approaches regarding the issue of the emergence of human behavior. Intended mainly as a guideline on how to construct scientific theories in anthropological research, the founding theoretical body presented here must be subjected to scrutiny by future research and “elucidated” both by new theoretical axioms and problem-testing processes. My prediction is that, if accepted, it will be modified, implemented, and better defined as expected by a philosophical perspective based on an evolutionary concept of knowledge.
Defining the concepts and formulating the hierarchy of the theory components
A theory requires a hierarchy of theoretical factual concepts, organized systematically. At the apex of a theory on the origins of human behavior is the generic ontological concept of “human behavior.” If we recall Isaac's (1978) diagnosis and expand it, we can structurally define human behavior, as opposed to nonhuman primate behavior, following the categories and components shown in Table 1.1. This reasoning is analogical and compares human and nonhuman primates, and the differences are the defining features. Such an analogical reasoning cannot be uncritically applied to the past, however. First, it is not articulated systematically within the body of a theory. Second, it does not define how many of these features are necessary to document in the evolutionary record to qualify as “human.” Third, it does not specify whether these features evolved independently or are interrelated. Fourth, no description was provided of how these features (converted into hypotheses) could be tested and what their falsifying premises would be. It is in these four lacking points that all strict analogical efforts applied to the interpretation of the archaeological record fail.
The formulation of a scientific hypothesis using Isaac's diagnostic criteria would imply placing human behavior within an evolutionary framework. Several theories could be used. For the sake of scientific development, I elaborate on one, acknowledging that alternative theories could be provided. The heuristics of those theories could potentially be confronted with the one defended in the present work, making up different research programs submitted to testing and subsequent differential explanatory power (Lakatos, Reference Lakatos1978). I am arguing that the origin of human behavior can be traced to the moment in which hominins shifted their subsistence toward a cooperative social organization, resulting in solidarity. Cooperative here means coordinated participation of all adult individuals in various subsistence activities. Solidarity refers to the end result of the expectation created by cooperation (Rankin and Taborsky, Reference Rankin and Taborsky2009), in which the outcome of any collective enterprise results in communal benefit. This should apply especially to the energy obtained through food and its relevance in adaptation. None of these features figures prominently in the behavior displayed by any extant nonhuman primate and currently is the basis of the structure of the behavior of our species (Quinlan, Reference Quinlan2008; Gurven and Hill, Reference Gurven and Hill2009).
A main theory requires, following traditional logic, a set of axioms postulated in the form of subsidiary smaller theories, also referred to as founder hypotheses (Bunge, Reference Bunge1998a). These serve as a starting point for deducing and inferring other subsidiary hypotheses that are made hierarchical relative to the founder hypotheses. As can be seen in Table 1.2, a set of axioms is presented as the pillar of the main theory. These axioms are not directly testable and must be supported by testable lower-hierarchy hypotheses, also referred to as intermediate- and lower-level theorems, and those that can be empirically tested in a direct way are referred to as factual hypotheses (Bunge, Reference Bunge1998a). The formal unity of a scientific theory consists of the existence of logical relations among the formulas of the theory, such that no formula remains isolated but remains fully integrated in a hypothetic-deductive body, which Bunge (1998a) referred to as the nervous system of a scientific theory.
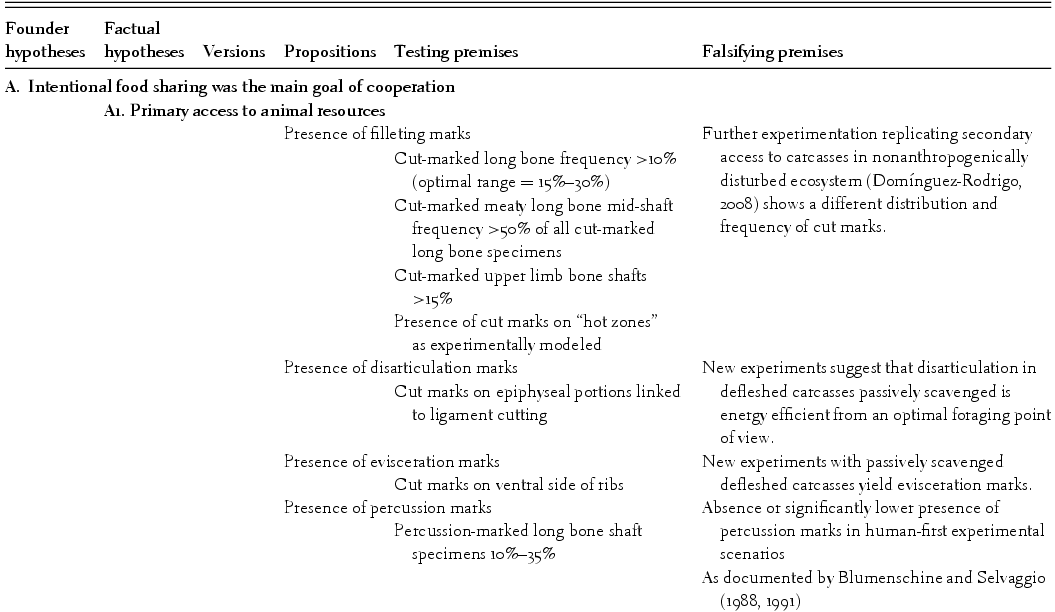
The axioms in Table 1.2 need to be defined and elucidated properly, that is, we need to sharpen their meaning to avoid confusion or ambiguity. This is a crucial step in the building of theories, because it provides meaning for each axiom and subsequently helps to make well-defined falsifying premises. The first axiom (intentional food sharing) refers to a subsistence strategy in which each individual belonging to a group forages with the intention of sharing an important part of the food obtained with other individuals within the reproductive unit and within the group. Ideally, the energy invested by each individual in obtaining food to be shared should account for a minimum of 30% to 50% of the total energy on which each individual's subsistence is based. This substantially differs from the food transfer documented among nonhuman primates (referred to by Isaac [1978] as tolerated scrounging), in which no energy is invested with the intention of obtaining food for others.
Food sharing requires food obtained in different places to be brought to a fixed place where it can be distributed. Hence the emergence of central places, which should be protective spots yielding safety for group fusion and where butchery and other activities might have taken place. I avoided the term home base, because the evidence of those spots acting as the focus of the remainder of social life and for sleeping remains archaeologically elusive. A referential place is a spot used by hominins for collective or individual activities other than food sharing that involves all members of the group or part of it, with the result of those activities yielding energetic benefit profited by more than one individual. These should be subordinated to central places. Several types of reference places can exist, which leave material traces that can be interpreted (see Tables 1.3 and 1.4).
A central element in food sharing among modern hunter-gatherers is meat (Stanford and Bunn, Reference Stanford and Bunn2001). It is therefore stressed that if food-sharing behaviors took place in the past, meat must have been a key element in them. For this purpose, meat consumption must have been a regular rather than a marginal (i.e., fallback) food type. By regular, it is meant that it was consumed year-round even if meat consumption peaked in certain moments (i.e., seasons) more than others.
Collective (cooperative) behaviors are a requirement prior to food sharing. They do not have to have been restricted to food obtainment and can be displayed in other activities. It is argued that the best way to trace them is to detect them in the obtainment and transport strategies of certain material elements that were required for the subsistence of hominins.
Our species is the only one on the planet whose survival and adaptability depends entirely on technology. It is also argued that one of our defining features involves not just use of tools in a more intense way than documented in other species (e.g., nonhuman primates) but also dependence on artifacts to the point where their removal from the behavioral repertoire would cause inadaptability. This idea of dependence might explain why stone artifacts appeared in human evolution. Archaeologists have been arguing for decades that stone tool use emerged in conjunction with meat eating as the essential butchering tools. Should meat eating be a marginal activity of hominins, then it should be inferred that stone tools were not used year-round and therefore that they were not essential to the survival of hominins. This suggests that this founder hypothesis should be studied in conjunction with the previous one.
Forethought and planning are described in terms of anticipation of future needs related to subsistence, reflected in the innovation of solutions prior to the appearance of the problem or need.
Finally, all of these axioms are conceived of within a systemic conception of behavior, as defended in the predominant behavioral ecology school of thought (Brooks and McLennan, 1991). This implies their interdependence and their emergence in a set, instead of each of them appearing at different times.
After each axiom has been defined as a founder hypothesis, which is the highest-ranked type of hypothesis, lower-ranked hypotheses stemming from the former are created to establish a link with the empirical record. These act as a bridge between the data and the founder hypotheses, which can be affected by various types of metaphysical components (and hence can be beyond direct testability). The factual hypotheses, because they concern facts and data (Bunge, Reference Bunge1998a), can be (1) analogical, when they emerge from the intuitive realization of similarities; (2) inductive, when they derive from examination of information; (3) intuitive, when their origin cannot be linked with either previous information or an analog; and (4) deductive, when they are derived from higher-ranked propositions. According to the degree of abstraction of a hypotheses, a further division can be made between observational and nonobservational hypotheses. The former are low-ranked hypotheses, because they are the first to make contact with reality through direct testing (factual hypotheses), whereas the latter are higher-ranked hypotheses, linking factual hypotheses to the main axioms of the theory. Hypotheses have also been described as phenomenological and representational, when addressing the external behavior of a theory without focusing on its working system and when they specify some mechanism, respectively (Bunge, Reference Bunge1998a).
Each of the factual hypotheses that make up the bulk of the theoretical body supported here, with some of their crucial testing and falsifying premises, can be seen in Table 1.3. The elucidation, sharpening, and definition of these hypotheses (senso Bunge [Reference Bunge1998a]) are described in Table 1.4. I will comment on only some of the reasons why some variables and hypotheses commonly used by some researchers to tackle the first founder hypothesis presented here were excluded. Food sharing requires food surplus, which in turn, if regularly obtained, requires primary access to carcass resources in modern African savannas. I have used some taphonomic indicators as analytical variables (converted into silogistic propositions inside hypotheses), which emphasize the careful and correct identification of hominin-imparted signatures preserved in the fossil record (namely, cut marks). This goes against a common trend among some Plio-Pleistocene archaeologists who postulate equifinality scenarios for those signatures and emphasize indirect reading of hominin authorship in any given bone assemblage through the imprints created by carnivores (Blumenschine, Reference Blumenschine1995; Capaldo, Reference Capaldo1995, 1997; Blumenschine et al., Reference Blumenschine, Peters, Capaldo, Andrews, Njau, Pobiner, Pickering, Schick and Toth2007; Ferraro, Reference Ferraro2007; Pobiner, Reference Pobiner2007). This may seem counterintuitive, and indeed it is. These authors claim that a high frequency of tooth-marked specimens on long bone midshafts is indicative of carnivores (namely, felids) having had priority in carcass exploitation and a significant input in the formation of bone assemblages, whereas a low frequency of tooth-marked specimens reflects the opposite, that is, the intervention of scavenging carnivores postdepositionally to ravage bones previously exploited by hominins. This assertion is incorrect for the following reasons, however: first, from a theoretical point of view it is aberrant, because to make the original claim of primary access by strict flesh-eating carnivores (felids, to enable hominins to intervene secondarily [as in Blumenschine, Reference Blumenschine1995]), experiments were conducted with durophage carnivores (e.g., bone-crunching hyenas) yielding high tooth-marking estimates, instead of with felids. Second, as expected, given the different tooth properties of felids and hyenids, taphonomic studies made on felid-eaten carcasses yielded much lower frequencies of tooth-marked broken bone specimens, indistinguishable from those abandoned by some durophage carnivores having secondary access to human-exploited bone assemblages (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007a, Reference Domínguez-Rodrigo, Pickering, Egeland, Pickering and Shick2007b). Researchers relying on carnivore-made tooth marks therefore should look for low frequencies of tooth-marked midshaft specimens if interpreting that flesh-eating carnivores had access to carcasses before hominins did. Third, tooth marks on midshaft portions of long bones are subject to equifinality: most researchers assume that they were caused by nonprimate carnivores, but nobody has been able to exclude other tooth-marking agents such as nonhuman primates (Pickering and Wallis, Reference Pickering and Wallis1997; Domínguez-Rodrigo, Reference Domínguez-Rodrigo and Ullrich1999), hominins (White and Toth, Reference White, Toth, Pickering, Schick and Toth2007), or suids (Domínguez-Solera and Domínguez-Rodrigo, Reference Domínguez-Solera and Domínguez-Rodrigo2009).
In contrast, the purported equifinality when using cut marks has been argued to be merely methodological; methods ignoring the real location of cut marks (linked to the behavior that caused them: filleting, dismembering, or skinning) and the type of bones where they occur yielded ambiguous interpretations (see extended discussion in Domínguez-Rodrigo [Reference Domínguez-Rodrigo, Hovers and Braun2009]), which can be overcome by using more accurate tallying methods (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007a). Furthermore, the use of “unknown” referents, such as other archaeological assemblages (which are assumed to be completely anthropogenic only because they are of late Pleistocene or Holocene age) is not only epistemologically wrong (an “unknown” cannot be used to explain another “unknown”) but taphonomically unjustifiable. Many of those Holocene sites used as analogs are either very fragmented diagenetically or have poorly preserved bone surfaces, which affects mark frequencies drastically. These recent sites cannot be directly compared to assemblages that lack diagenetic breakage and have well-preserved cortical surfaces.1 Domínguez-Rodrigo and Yravedra (Reference Domínguez-Rodrigo and Yravedra2009) showed that variability in percentages of cut-marked bones is tightly related to several different variables, but when cortical preservation is controlled, there is an expected range of cut-marked specimens that enables the use of cut-mark frequencies as a good indicator of the anthropic component of any given assemblage.2 Ideally, comparisons among assemblages have to be made when it has been confirmed that all the variables involved are equally (un)biased. All the taphonomic components of the hypotheses contained in the theory that I present affecting bone surfaces are based on well-preserved assemblages and should be applied as analogs on the unbiased parts of archaeological assemblages; that is, the sample comprising specimens with well-preserved cortical surfaces.
The types of factual hypotheses proposed for the theoretical body presented here are observational (ten factual hypotheses) and nonobservational (seven axiomatic hypotheses; Figure 1.1). The factual hypotheses are either deductive (stemming from hypothetic-deductive inferences; A2, B2, D1, D2, F, G) or deductive-analogical (A1, B1, C1, E). Strict analogical hypotheses have not been considered.
Hypothesis interrelatedness: The neural network of the theoretical body
Processual approaches to anthropological issues have provided hypothesis-testing frameworks, often testing opposite propositions beyond the null hypothesis. The most common pattern for anthropologists is to test hypotheses individually, however. This limits the heuristic capacity of each hypothesis given the common phenomenon of equifinality in historical and contextual processes. By linking hypotheses, this problem is frequently overcome, given that the heuristic value of each hypothesis is reinforced by those of the other hypotheses to which it is linked, and therefore on which it is dependent. For instance, the hypothesis that hominins focused on the exploitation of large carcasses (especially those larger than 100 kg; Hypothesis A2) is constructed on the premise that accumulations of animals these sizes must occur at sites. Taken in isolation, this assumption invalidates the hypothesis because other nonanthropogenic processes (e.g., natural deaths at ponds during droughts, accumulation of carcasses by certain carnivores in dens or death arenas) can produce the same result. If it is linked to hypothesis A1, the heuristic value of hypothesis A2 increases, because the co-occurrence of an accumulation of carcasses of that size range on the same spot with unambiguous evidence of having been primarily exploited by hominins invalidates the purported equifinality with a scenario of carcasses accumulated by carnivores (and initially consumed by them). If the link of A2 is extended to B1 (selection of central places), the other possible equifinality scenario of hominins exploiting naturally accumulated carcasses by catastrophic phenomena vanishes, because B1 requires taphonomic support for dynamic (transport-caused) processes in carcass accumulation.
The relationship between linked hypotheses can be of reciprocity (one hypothesis and another show similar degrees of dependence), or of asymmetry (one hypothesis is unidirectionally dependent on another without corresponding dependence on it). The linking can be double or multiple, involving several related hypotheses (Figure 1.1).
A neural network conception of hypotheses shows that the testing of independent hypotheses could be futile, because no hypothesis ever exists independently within a theory. Independent hypothesis testing is good for discarding falsified hypotheses but inappropriate for selecting among corroborated hypotheses (those prone to create equifinality scenarios). Lyman (Reference Lyman2004) correctly stressed that equifinality in open systems was a methodological construct.3 Equifinality in such systems is not an inevitable situation. Current methods can be inadequate to differentiate causes from their effects, but alternative ones could potentially do so, because natural historical processes are open systems (Lyman, Reference Lyman2004). This conception applies perfectly to hypothesis testing. When two hypotheses have been corroborated (which should not be mistaken for confirmed) by testing, the only way to select between them is to provide different testing premises or methods, or as scientific realists advocate, to construct meta-hypotheses, which involve lumping the propositions of two originally separate but dependent (theoretically linked) hypotheses into one broader hypothesis (Bunge, Reference Bunge1998a). This increases the discriminatory power of testing.
The corroboration of single hypotheses also potentially increases the value of other hypotheses to which those are linked. The dendritic relationship of conceptual interdependence of hypotheses determines the heuristic value of theories.
In the theory constructed here on the origins of human behavior, most factual hypotheses show a direct link to the factual hypotheses from which they derive and also single or multiple dependence links to other hypotheses, as can be seen in Figure 1.1. This link is explained in Table 1.4. This theory can be used as a good demonstration that the contribution of each hypothesis to the main axiom of the theory is not even. Some hypotheses contribute more crucially than others. For instance, A1 shows mostly unidirectional links to almost one half of the other hypotheses, which indicates that 45% (five of eleven links) of the heuristic power of the theory depends on A1; that is, the extent to which high resolution can be reached in discerning primary access by hominins to large carcass faunal resources, producing the food surplus necessary to justify intentional food sharing by its repeated transport to the same locus.
How to measure the heuristic value of alternative theories?
Lakatos (Reference Lakatos1978) argued that programs of scientific research compete with one another according to their heuristic value, that is, their capability of explaining a bigger portion of reality. For those who argue that refutability is never absolute, the selection of hypotheses and theories should be made based on their capability of explaining things. Lakatos argued that research programs were endowed with auxiliary hypotheses (a theory's protective belt) that rendered falsification less straightforward than a Popperian approach, focusing on refutability, would suggest. It is therefore the capacity of interpreting reality by explaining things that discriminates among hypotheses. Lakatos suggested that rather than selecting one program over another because one is refuted, selection is made because one program will always discover novel facts, showing continuous growth. This qualifies as a progressive research program in opposition to a degenerative research program, which is marked by lack of growth owing to the effect of the protective belt of auxiliary hypotheses. The degeneration of a program causes its falsification (Lakato's sophisticated falsifiability).
If we switch “research program” to “theory,” then it is clear that irrespective of the degree of refutability of a theory's components (hypotheses) or lack thereof, the degree of corroboration and the amount of knowledge conveyed by alternative theories determines which one is epistemically selected and which one vanishes. It is clear that progressive theories, which contribute with more knowledge, will be preferred to those that are regressive or degenerative. The degree of heuristic value of any given theory is perceived when the differences are large, however. In theories with a smaller degree of heuristic difference, this perception is less obvious.
Scientists should ideally dispose of a “heuristometer,” which could provide objective value of the explanatory power of a theory, but that is missing in nonapplied sciences. For historical sciences, in which the quantification of the amount of corroborated knowledge provided is less certain than for applied experimental sciences, a theoretical indicator should be created. Figure 1.2 suggests an example of one of the many plausible indicators for differentiating the explanatory power of scientific theories created in anthropological research.
The amount of empirically contrasted knowledge contained in a theory is directly related to the number of factual hypotheses contained in it. Therefore, a theory with a greater number of hypotheses is more difficult to corroborate than another theory with a smaller number of them, although if corroborated even partially, the former could provide more information than a smaller theory, in which all the factual hypotheses have been corroborated. For this reason, any measure of explanatory power must take this into account. Each hypothesis therefore represents a proportion of potential factual knowledge, which varies in each theory according to its size. Furthermore, each hypothesis within a theory does not contain the same amount of empirically tested information, but it varies according to how relevant or peripheral the hypothesis is. The degree of relevance is marked by the number of dependent relations that the hypothesis holds with other hypotheses within the theoretical body (i.e., number of links). To establish a reliable indicator of the amount of knowledge contained in any given theory, the number of hypotheses contained and their types of dependence among one another should be considered.
This would allow one to estimate the relative amount of corroborated proportional knowledge, referred to as the number of hypotheses that have been successfully tested, derived as a relative proportion of values obtained when considering the number of hypotheses involved and their value in the theoretical body. It would also allow one to estimate the potential amount of noncorroborated proportional knowledge, which refers to the number of hypotheses that either have not been successfully tested yet or that have been empirically rejected. Both the corroborated set of hypotheses and the noncorroborated set of hypotheses constitute the total potential factual knowledge of a theory. Using these parameters, as shown in Figure 1.2, can lead to a primitive indicator of the explanatory power of a theory, or heuristic index.
To apply this index accurately, one should consider whether both theories tested share the same question (in this case: what is the origin of human behavior?) even if the main axiom (e.g., cooperative behavior leading to solidarity or, alternatively, any other proposition) differs in each theory.
Discussion and conclusion
I do not intend here to provide arguments why a processual approach to anthropology is more accurate or advantageous than other theoretical approaches in terms of acquisition of knowledge. Rather, the main goal of this text is to create awareness among scientific anthropologists that their procedures might or might not qualify as scientific, depending on the philosophical school of thought that they conscious or unconsciously embrace. Today the dominant epistemological school for practitioners of natural sciences is scientific realism.
Almost all postmodern approaches to anthropology see the scientific effort of the processualist approach with skepticism. Beyond their constructivist vision of science, their view is fueled by the unfinished debates created within the processual approach itself. A purported scientific hypothesis frequently leads to interpretations that are widely divergent from another “scientific” hypothesis also aiming at testing the same question. My humble view is that part of the explanation for this is the incompleteness with which several processualists have understood the relationships between theory-hypothesis and axiom-theorem (Bunge, Reference Bunge1998a). Processualists began constructing their conceptual and theoretical building correctly but stopped once the foundations of the building (how to create factual hypotheses and empirically test them one by one) were finished. They also know how to build the roof (the theory containing the main axiom), but they are missing the bulk of the building (structural frame and walls) because they mostly have not articulated hypotheses successfully and hierarchically (from factual to founder hypotheses or vice versa), linking them conceptually to the theory in an interrelated way (most hypotheses of a theory are interdependent or systemic; Bunge, Reference Bunge1998a, 2006). This criticism is valid only if it is assumed that to gain parity with natural sciences, scientific anthropology is better defended from a scientific realistic epistemic approach. The denial of this assumption, as well as the refusal to follow scientific methods, would lead to the rejection of the bulk of the ideas expressed in this work.
Scientific realists conceive theories as permanently morphing through the testing of their hypothesis (Figure 1.3) because of an increasing body of knowledge. In the words of Bunge (Reference Bunge1998a, p. 436)
Since the data-gathering-and-packaging view of science ignores the aims of theorizing, it will be convenient to state such aims explicitly. The basic desiderata of scientific theory construction are the following. (i) To systematize knowledge by establishing logical relations among previously disconnected items; in particular, to explain empirical generalizations by deriving them from higher-level hypotheses. (ii) To explain facts by means of systems of hypotheses entailing the propositions that express the facts concerned. (iii) To increase knowledge by deriving new propositions (e.g., predictions) from the premises in conjunction with relevant information. (iv) To enhance the testability of the hypotheses, by subjecting each of them to the control of the other hypotheses in the system.
The latter part of this statement is crucial in the way factual hypotheses are tested and has been regularly neglected in “scientific” approaches to anthropology. This explains in part the frequent low resolution of processual projects when displaying contradictory interpretations, despite employing the empirical-hypothesis testing approach.
Factual theories are iconic reconstructions of real systems and are inferred models of reality. Modeling is an essential part of theory building, and most factual hypotheses in anthropological theory are analogical; however, this must not be mistaken with the use of analogies as proxies for theories. In the field of early human evolution, the reconstructions of hominin behavior from the Plio-Pleistocene record have been based on pure analogical reasoning. Isaac (Reference Isaac1978) interpreted sites by using an analogically derived list of characteristics in which human and nonhuman primates differ. Binford's (1981) subsequent criticism, although better framed from a theoretical point of view, was also based on a structured and limited view of analogy (through the middle-range theory), by comparing a varied repertoire of analogies to published data from early sites: do sites look like human foragers’ base camps or carnivore-formed assemblages? Binford contributed positively when looking at alternative possibilities to the anthropogenic models widely held, which could fit the empirical record better. His “marginal scavenger” model was produced as an ad-hoc interpretation to statistical treatment of data, which did not fit quite well with either strict human or carnivore models of bone accumulation. Paradoxically to his hypothetic-deductive philosophy, it also did not stem from a theory conceived prior to empirical testing but rather was inductively built a posteriori without the support of a scientifically framed hypothesis specifying what a bone accumulation modified by a marginal scavenger hominin must look like. Sept's [1992] chimpanzee-nesting model was another example of analogical interpretation not epistemologically justified: just because chimps redundantly occupy the same trees for nesting, hominins could have done something similar to generate the characteristics observed in the early archaeological record. Rose and Marshall's (1996) “resource-defense” model also failed to be theoretically framed in a similarly scientific way, with the elaboration of testable hypotheses, and was mainly based on ethological analogy. Blumenschine's (1986) passive scavenger model was also constructed by mere ethological/ecological analogy of observed availability of carcass resources in one ecosystem assuming the same processes could have existed in the past (without contemplating variability caused by different ecological conditions [see Tappen, Reference Tappen1992, Reference Tappen1995]). He initially did not produce a scientific hypothesis with clearly defined propositions and falsifying premises. When he did (Blumenschine, Reference Blumenschine1995), these were subsequently refuted (review in Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007a). O’Connell's (Reference O’Connell1997) near-kill location model also failed to be phrased in the form of a testable theory and was based on descriptive ethnographic analogy: if modern foragers create bone accumulation near alluvial environments produced by their preparation of carcass remains to be transported elsewhere, why not assume that early sites had a similar function?
Ever since the revisionist debate started in the 1980s, the different behavioral models created to explain the early archaeological record have been either strictly analogical (most models), purely theoretical without properly phrased testable hypotheses (e.g., Pott's [1988] stone-cache model4), or completely lacking hypotheses and a scientific theoretical body (e.g., the preferred site model [Schick, Reference Schick1987] or the male display model [O’Connell et al., Reference O’Connell, Hawkes, Lupo and Blurton-Jones2002]). Less frequently, when some authors provided a better-described testable hypothesis (passive scavenging in barren floodplain and opportunistic refuge [Blumenschine and Masao, Reference Blumenschine and Masao1991; Blumenschine et al., Reference Blumenschine, Cavallo and Capaldo1994]), this could be tested and subsequently refuted (Ashley et al., Reference Ashley, Domínguez-Rodrigo, Bunn, Mabulla and Díez-Martín2010), enabling some advance in our understanding of early human behavior. The choice of models available in recent and not so recent literature is so long that modern archaeologists are confused as to (1) which one fits the empirical record better and, (2) whether uncovering the behavioral meaning of early sites is theoretically feasible. Thus, is it not strange that recent so-called behavioral models have given up tackling the functionality of sites and focus on the hominin subsistence strategy that could be most directly read from the materials contained in sites (e.g., Ferraro's [Reference Ferraro2007] obligate carnivory model).
It could be argued that such a variety of interpretations (most of them stemming from processual approaches to the archaeological record) reflects (1) intensive and abusive use of analogy expressed either through direct application of the analogue to the archaeological record, without a properly defined theory or (less ambitiously) a hypothesis; (2) individual hypothesis testing without framing it in a systemic theoretical body; and (3) lack of links among hypotheses, resulting in insufficient testing for resolving equifinality scenarios and opposite interpretations. Processualists might believe that the empirical testing of individual hypotheses is scientific enough, but by doing this the hypotheses thus tested are devoid of a substantial part of their heuristic power: the one generated by its association with other hypotheses.
I would argue that part of the reason why processualists specialized in testing single hypotheses derives from the application of Binford's “middle-range theory” approach. Middle-range theory in archaeology was borrowed from its counterpart in sociology, as pointed out earlier. Merton (Reference Merton1967) had argued that to adapt theory to its empirical consequences, scholars had to give up the attempts of finding a general theory that would explain holistically all aspects of social life. A “grand theory” would be hard to test empirically, especially in a field that lacked control of most of the variables, such as sociology. Merton argued that sociologists had to concentrate on fragments of social reality until the body of knowledge acquired would enable sociology to converge with natural sciences and provide explanations in the form of laws or patterns of human social behavior. Theories, in their broadest sense, had to be avoided (but only initially). Binford (Reference Binford1981) borrowed the approach literally by linking middle-range theory to determined aspects of human behavior or natural processes. Middle-range theory in archaeology provides answers to specific functional questions; that is, how a determined process results in certain material diagnostic characteristics (e.g., how certain butchery practices leave diagnostic features on bones). By deliberately emphasizing the role of middle-range theory, Binford avoided the testing of larger theories. Middle-range research was conceived as a prerequisite to the enquiring of questions that implied a combination of more than one process. It would not be an exaggeration, however, to say that the role that middle-range theory played in processual archaeology was the same as that of scientific factual hypotheses in applied sciences. It limits testing to specific hypotheses. Processualists have not realized that a systemic use of middle-range theory would enable them to test grand theories only if processes could be intertwined. Linked hypotheses provide higher heuristic power, which would make their testing through middle-range approaches more efficient and discriminatory. Instead of that, Binford's limited conception of the use of middle-range theory does not go as far as Merton's, which implies that when the knowledge provided by middle-range theories builds, then grand theories can be approached. This handicaps Binford's goal of making archaeology a fully scientific endeavor, because isolated hypotheses cannot be efficiently patched to construct reality (Bunge, Reference Bunge1998a).
In Plio-Pleistocene archaeology, this translates into one basic consequence: the issues of the emergence of stone tool use or meat eating are not independent from the fact that both behaviors occur in selected spots where materials accumulate in high densities, which archaeologists call sites. Something as complex as the meaning of early sites cannot be grasped through middle-range theory alone but through a wider range theory. Stone tool use or meat eating, thus, cannot be explained independently from the behavioral meaning of those sites where they occur. Archaeologists have been blinded by the immediacy of the middle-range theory. It applies to factual hypotheses alone, but its power would be broader if applied systemically. The time is ripe for processualists to think bigger; grand theories (as in natural sciences) should guide the way of scientific research in anthropology, knowing that the building of scientific knowledge implies starting empirical testing by the foundations (factual low-level hypotheses) and going upward toward the roof (founder hypotheses and main axiom of the theory).
The goal of this work is to present the structure of a scientific theory on the emergence of human behavior without putting it to test. That would take another significantly longer work. I can predict, however, that the status quo of similar heuristic value or ambivalence affecting most of the available models to explain early archaeological sites will vanish when this, or other similarly framed theories, are applied to the fossil record. I also predict that most of these models will be definitively discarded.
Table 1.1. Principal elements for a definition of human behavior (drawn from comparison with nonhuman primates)

Table 1.2. A proposed theory for the origin of human behavior









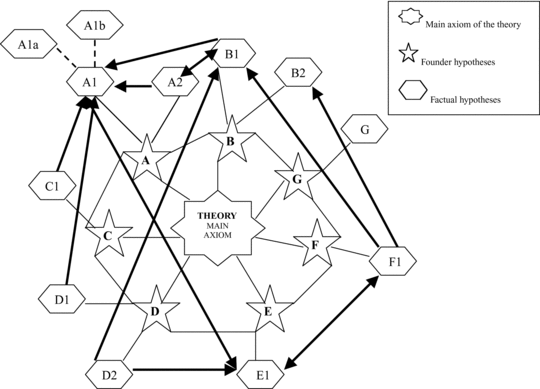
Figure 1.1. Interrelations among founder and factual hypotheses, with special emphasis on the links among different factual hypotheses (thicker lines). Direction of interdependence between hypotheses is indicated by arrows. See key for each hypothesis in Tables 1.2 and 1.3.

Figure 1.2. An example of a suggested heuristic index (H.i; see text for explanation) comparing its utility with the theory proposed in this paper (A), assuming that only a portion of hypotheses have been successfully empirically supported (A1, A2, B1, B2) and two alternative theories, in this case consisting of the same theory but containing a smaller number of successfully tested hypotheses (A1,A2); (B) or a smaller theory containing a smaller number of factual hypotheses, with a similar proportion to A of those empirically corroborated (A1, A2, D2); (C) The value of the heuristic index is expressed in absolute numbers; the higher the result the more explanatory power the theory contains. In this case, A (0.088) contains a higher amount of information explained and empricially corroborated than B (−1.14) or C (−0.27). C, despite being a smaller theory and containing a smaller explanatory potential, has a higher explanatory power than B because in the latter only two hypotheses were empirically corroborated. A negative index shows a negative balance of the potential amount of explanatory power contained in the theory, when comparing the hypotheses that have been corroborated and those remaining to be empirically corroborated. Heuristic index created using R (http://www.r-project.org/).
References
1 This, for instance, happens when using assemblages like SK400 (LSA, South Africa; Dewar et al., Reference Dewar, Halkett, Hart, Orton and Sealy2006) as referents of low frequencies of cut-marked bone (<1%) in anthropogenic assemblages (for instance, Ferraro, Reference Ferraro2007), when almost one-third of the bones are burnt; the remainder seem to be fairly badly preserved, covered with root etching and sediment (Dewar et al. [2006], figure 5); and bones were not clean when their surfaces were analyzed for marks.
2 Domínguez-Rodrigo and Yravedra's (Reference Domínguez-Rodrigo and Yravedra2009) comprehensive review of cut-mark frequencies in twenty-eight sites, including forty-four archaeological levels spanning different chronologies, based on the well-preserved green-fractured assemblages, excluding those specimens with poor cortical preservation, yielded an average of cut-marked long bone specimens of 19.9% (median = 19), standard deviation of 11, with 80% of the sample showing the following percentile (10–90) range of cut-marked long bone specimens: 6%–36%.
3 Lyman (Reference Lyman2004) urged readers to look at equifinality from its original meaning as defined by von Bertalanffy (Reference von Bertalanffy1956) when modeling general systems theory, that is, the “same final state from different initial states” in an open system.
4 Actually, critics of the model were responsible for empirically testing it and providing falsifying scenarios (de la Torre and Mora, Reference Torre and Mora2005; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007a).