

The Atlantic Sea Scallop fishery has experienced dramatic increases in biomass, landings, and prices over a twenty-year time period (Hart and Rago Reference Hart and Rago2006, Northeast Fisheries Science Center 2014). We use a set of econometric methods and models to understand how port-level landings have responded to changes in proximity to biomass. Access to biomass, a particular type of natural advantage, fluctuates from year-to-year in this fishery and may cause long-lasting changes in the geographic distribution of fishing as agents respond to changing environmental and regulatory conditions by investing in capital or relocating existing capital (Portman, Jin, and Thunberg Reference Portman, Jin and Thunberg2009, Reference Portman, Jin and Thunberg2011). In addition, within- and across-fishery agglomerative forces (separate from any natural advantages conferred by access to biomass) may influence where scallops are landed as participants in the industry respond to economies of scale and scope. We examine these effects by estimating long-run economic growth models that relate growth of scallop fishing over fifteen years to initial conditions and exposure to biomass. Fluctuations in natural and regulatory conditions may also have effects that vary across ports or occur at a faster time scale. We test for the existence and relative magnitude of these effects using time-series methods.
Fisheries management in the United States has many goals; these include mandates to prevent overfishing, provide the greatest overall benefit to the nation, and take into account the importance of fishery resources to fishing communities (Magnuson-Stevens Fishery Conservation and Management Act 2007). The effects of natural advantages relative to endogenously determined capital has implications for resource managers concerned with fishing communities. If port-level landings are very sensitive to biomass, then ports will quickly expand or contract after experiencing temporary biomass shocks. However, if landings are not sensitive to biomass, then it may be necessary to identify ports that are declining, and understand why they are doing so, prior to formulating an appropriate policy response designed to address fishing communities.
Furthermore, economic studies of cities illustrate that changes in economic activity in one industry could have secondary effects on other parts of the local economy through spillovers (Quigley Reference Quigley1998, Puga Reference Puga2010, Behrens, Duranton, and Robert-Nicoud Reference Behrens, Duranton and Robert-Nicoud2014). The commercial fishing industry is linked to upstream and downstream industries, including seafood processors, wholesalers, gear suppliers, settlement houses, and repair operations (Kaplan Reference Kaplan1999, Hall-Arber et al. Reference Hall-Arber, Dyer, Poggie, Mcnally and Gagne2001, National Marine Fisheries Service 2016). The potential for spillover effects on other fisheries and related industries has induced local governments to develop port management plans (Georgianna et al. Reference Georgianna, Avila, Bethoney, Cass, Challingsworth and Hogan2014) or directly lobby the executive and legislative branches of the U.S. government to preserve local fishing capacity (Bonner Reference Bonner2017, Valencia Reference Valencia2017).
Marshall (Reference Marshall1920) posited natural advantages, economies of scale, input sharing technology adoption, and labor market matching as possible drivers for clustering and agglomeration of economic activity. Re-energized by Krugman (Reference Krugman1991), a substantial burst of research has been directed at characterizing the agglomeration of economic activity at fine scales and understanding which of these Marshallian forces is most importantFootnote 1. Kim (Reference Kim1999), Ellison and Glaeser (Reference Ellison and Glaeser1999), Midelfart-Knarvik et al. (Reference Midelfart-Knarvik, Overman, Venables and Helene2001), Bleakley and Lin (Reference Bleakley and Lin2012) find support for the importance of factor endowments in U.S. manufacturing location, U.S. manufacturing agglomeration, EU industrial location, and U.S. economic activity respectivelyFootnote 2.
Extensive research has demonstrated the importance of natural advantages, as measured by high catch rates or low costs, in determining where fishing activity takes place (Girardin et al. Reference Girardin, Hamon, Pinnegar, Poos, Thebaud, Tidd, Vermard and Marchal2017). Quite a bit less is known about ports. Portman, Jin, and Thunberg (Reference Portman, Jin and Thunberg2009, Reference Portman, Jin and Thunberg2011) show that healthy biomass signals high future profits and encourage development of land to support the fishing industry. Watson and Johnson (Reference Watson and Johnson2012) finds evidence that nonspatial policies can have heterogeneous effects across the coast as well. Watson and Johnson (Reference Watson and Johnson2012) examine changes in landings on the West Coast of the United States after a fishery buyback in 2003 using static panel methods, finding that small ports had higher variability in landings after a buyback than large ports. Speir, Pomeroy, and Sutinen (Reference Speir, Pomeroy and Sutinen2014) and Lee et al. (Reference Lee, Speir, Carr-Harris and Benjamin2017) characterize but do not explain spatial distribution of fishing for fisheries on the East and West Coast of the United States. Agnarsson, Matthiasson, and Giry (Reference Agnarsson, Matthiasson and Giry2016) examine distribution of quota across ports in Iceland, which is likely to be closely related to landing locations.
Both first-nature (natural advantages) and second-nature (caused by humans) geography are likely to be important. For example, ports that are located near productive fishing locations will have first-nature geographic advantages relative to ports that are farther from good fishing grounds. Similarly, endowments of social (Kaplan Reference Kaplan1999, Hall-Arber et al. Reference Hall-Arber, Dyer, Poggie, Mcnally and Gagne2001, Holland, Pinto da Silva, and Kitts Reference Holland, Pinto da Silva and Kitts2015, Clay, Colburn, and Seara Reference Clay, Colburn and Seara2016) and physical capital can confer second-nature advantages to established ports to nontraditional fishing ports. The Atlantic Sea Scallop fishery provides a unique opportunity to study the importance of natural advantages relative to “second-nature” geography. Unlike finfish, scallops are relatively immobile after settling on the ocean floor (Hart and Rago Reference Hart and Rago2006). There is extensive fishery-independent survey data that provides information about the location and scale of biomass. The geographic distribution of fishable scallop biomass has also changed frequently due to both natural variability and area-based fishing regulations (Hart and Rago Reference Hart and Rago2006). The management system, which opens and closes areas of the ocean to solve the growth overfishing problemFootnote 3, can dramatically change a port's access to biomass from one year to the next.
We use two sets of methods to examine the effects of changes in natural advantages on ports and fishing activity. We first estimate a long-run growth model to examine how natural advantages (access to biomass and variability of access over time) and returns to both scale and scope influence scallop landings at the port level. We also estimate time series models that explore the relationship between natural advantages and landings at a finer temporal step for the twenty-five largest scallop ports in the region.
Scallops, the Fishery, and Regulations
The Atlantic sea scallop is found on the continental shelf of the Northeast United States, from North Carolina through Maine. Important fishing grounds are on Georges Bank (GB), Southern New England (SNE), and the Mid-Atlantic Bight (MAB), at depths up to about 350 feet (Hart and Chute Reference Hart and Chute2004, Hart and Rago Reference Hart and Rago2006). Scallops reproduce by producing large amounts of eggs; larvae subsequently drift with water currents before settling to the bottom of the ocean (Hart and Chute Reference Hart and Chute2004). Under favorable conditions, this can result in extremely high abundances of juvenile scallops in localized areas. The biological characteristics of sea scallops make them particularly well suited to spatial management: scallops grow relatively quickly, adults typically have low natural mortality, and scallops are relatively immobile after settling on the ocean floor (Hart and Rago Reference Hart and Rago2006). The biomass of scallops is currently high (Northeast Fisheries Science Center 2014). Scallops in GB and MAB waters grow similarly until they are about 8 cm shell height, but scallops on GB grow to a larger asymptotic size (Hart and Chute Reference Hart and Chute2009). In both areas, scallops grow to a smaller asymptotic size in deeper depths; this effect is more pronounced in the MAB (Hart and Chute Reference Hart and Chute2009).Footnote 4
The National Marine Fisheries Service (NMFS) is responsible for assessing the sea scallop population and fishery in federal waters off the coast of the US; this has historically been accomplished using a dredge survey and commercial data. Beginning in the mid-2000s, this was supplemented by a pair of optical surveys that increased the intensity and geographic breadth of scientific information. As a result of the high intensity of survey efforts, the biomass and location of scallops are quite well understood.
The New England Fishery Management Council, composed of industry stakeholders, non-industry stakeholders, state officials, and federal officials, is responsible for recommending scallop fishery policy in federal waters (3–200 nautical miles from shore). NMFS translates these policies into regulations and enforces those regulations. Fisheries policies are guided primarily by the Magnuson-Stevens Fishery Conservation and Management Act, which describes ten standards for fisheries management. The charge to prevent overfishing and achieve “optimum yield” (Magnuson-Stevens Fishery Conservation and Management Act 2007) is frequently the most difficult to achieve; however, the MSFCMA also requires understanding how regulations affect fishing communities and accounting for those effects when setting policy. Fishing vessels in the Atlantic sea scallop fishery typically use dredge gear to harvest scallops from the ocean bottom Northeast Fisheries Science Center (2014). Processing of scallops, in which the adductor muscle is retained and sorted by size, has occurred on-board the vessel since the mid 1930s (Georgianna, Lee, and Walden Reference Georgianna, Lee and Walden2017). As part of the fishery management process, NMFS requires both fishing vessels and fish buyers to obtain permits. Fishing vessels are required to file trip reports, and buyers are required to report purchases of scallop from those fishing vessels. Because the primary objective of these data collection efforts are to determine the amount of fish and shellfish that are harvested, there are no reporting requirements for subsequent sales further up the supply chain.
In the scallop fishery, fishing regulations primarily include limits on days at sea (DAS), gear restrictions, crew restrictions, and temporary closures of parts of the ocean, so that the annual catch limits (ACLs) are not exceededFootnote 5. Since 1994, scallop vessels were allocated a number of DAS which they could use in “open” areas. Spatial management has featured prominently in scallop management since early 1994, when two large areas on Georges Bank (Closed Area I and II) and another in Southern New England (Nantucket Lightship) were closed to bottom-tending gear, including dredges and bottom trawls, to rebuild depleted stocks of groundfish (59 Federal Register 26; 59 Federal Register 9872). Two areas in the Mid-Atlantic were closed in 1998 to allow juvenile scallops to mature. The scallop fleet was allowed to fish in a portion of Closed Area II in 1999 and the access program was expanded to parts of Closed Area I and Nantucket Lightship in 2000. The Mid-Atlantic areas were reopened as rotational access areas in 2001. Vessels fishing in these areas were required to use DAS and were subject to a possession limit (typically a trip “cost” 10 DAS).
In 2004 the current management system, consisting of access, open, and closed areas was adopted (69 Federal Register 35194–224). One of the southern access areas (Virginia Beach) was reconfigured as an open area because it was not successful at encouraging growth of scallops. Additional areas have been added to the rotational program; the boundaries of these areas experienced minimal changes until 2015, when the Mid-Atlantic regions were reconfigured into a single large access area and a small closed area. In the current system, an area can be closed to scallop fishing when it contains high abundances of juvenile scallops. This allows scallops to grow larger, addressing the growth overfishing problem. When opened, fishing vessels are allocated trips, with a possession limit, into the access areas. These trips no longer require using open-area DAS.Footnote 6
As access areas are opened and closed, and biomass in all parts of the ocean fluctuate, the relative advantages of fishing from a particular port are likely to change. Increases in travel time to the rotational areas leads to increased expenditures on fuel. Increases in travel time to open areas are even costlier because fishing vessels also must use scarce DAS to steam instead of actively fish. Fishing vessels are quite mobile and can respond to spatial variations in costs by changing locations. It seems natural, then, to examine the effects of changing resource advantages on ports.
Data
The subsequent analysis primarily relies on two datasets containing data from 1996–2015. Exploitable biomass in each of the twelve discrete areas was constructed using fishery independent data collected through dredge and optical surveys. These estimates were aggregated to construct an available biomass index using the schedule of openings and closing taken from the fishery regulations published in the Federal Register. The second dataset contains information on landings and value, by species, aggregated to the port level. The major ports and biomass regions are illustrated in Figure 1.

Figure 1. Study Area with Biomass Regions and Largest Twenty-Five Ports
We constructed an inverse-distance weighted biomass availability index to measure natural advantages in each port. We used estimates of exploitable biomass density (kilograms per standardized tow, only individuals larger than 90 mm) in each of twelve discrete areas where scallops are harvested (Figure 2)Footnote 7. Each of the twelve biomass regions was decomposed into small (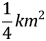 $ \displaystyle{1 \over 4}km^2 $) cells. We index ports, fishing region, and cells with i, r, and k subscripts respectively and suppress the time subscript for compactness. Each fishing region has K r cells. Ordering the regions so that the regions indexed r = 1, …R 1 are open and the rest are closed, the available biomass index (Available i) is:
$ \displaystyle{1 \over 4}km^2 $) cells. We index ports, fishing region, and cells with i, r, and k subscripts respectively and suppress the time subscript for compactness. Each fishing region has K r cells. Ordering the regions so that the regions indexed r = 1, …R 1 are open and the rest are closed, the available biomass index (Available i) is:
 $$Available_i = \sum\limits_{r = 1}^{R_1} {\sum\limits_{k = 1}^{K_r} {\displaystyle{{Exploitable\; Biomass\; Density_r} \over {Distance_{ik}}}}}\comma \; $$
$$Available_i = \sum\limits_{r = 1}^{R_1} {\sum\limits_{k = 1}^{K_r} {\displaystyle{{Exploitable\; Biomass\; Density_r} \over {Distance_{ik}}}}}\comma \; $$where Distanceik is the over-water travel distance (kilometers) from port i to cell k. This formulation allows the available biomass index to vary both across ports within a year due to different distances, and across years within a port as stock levels fluctuate and areas of the ocean are opened and closed. It also is constructed so that differently sized regions contribute in proportion to their size.

Figure 2. Exploitable Biomass Density in the Twelve Areas. Years in Which a Region was Closed to Scallop Fishing are Shaded in Gray
The available biomass index is plotted for four ports to illustrate the variability in this index: New Bedford, MA (north); East Hampton, NY (central); Cape May, NJ (south); and Newport News, VA (far south) in the upper panel of Figure 3. New Bedford, MA is located at approximately the same latitude as the northernmost scalloping area in Georges Bank, while Newport News, VA is slightly south of the southernmost scalloping area in the MAB. There is variability both within- and between ports. The lower panel of Figure 3 illustrates the rank order of the biomass index for these four ports. There is a substantial variability in this measure of resource advantage. This variability is driven by both large, localized recruitment events and by openings and closings of areas by managers. However, most of the ports that are persistently ranked lowest are in Virginia, which is the southernmost portion of the scallop habitat.

Figure 3. The Available Biomass Indices (Left) and Rank (Right) for Four Selected Ports in the Northeast United States
Landings and value are constructed using vessel trip report and dealer report data. The vessel trip report data have been mandatory for federally permitted fishing vessels since 1994; however, the first two years of data have many data errors, only data from 1996–2015 were used. The vessel trip report data are used as the source for landings (quantities) and port landedFootnote 8. The dealer data are used to construct prices needed to compute value. We aggregate to the 2013 version of the U.S. Census county subdivision to construct a dataset of annual landings, based on the scallop fishing year, and value for each species. While vessel captains report the port of landing, the precision of this particular data field seems to have varied across captains and over time. The county subdivision strikes a reasonable balance between high spatial resolution and potential error in classification. Fishing vessels have landed scallops in 177 distinct ports in the northeast U.S. over the past 20 years. However, the largest 25 ports account for 98.6 percent of the fishery value over the 20-year time period. Among the top three to four ports, there has been minimal fluctuation in the rank order of scallop landings; however, there has more variation for the medium and small scallop ports (Figure 4). While we would ideally include controls for the location and scale of scallop processing firms in our econometric analysis; the data collection protocol for NMFS's annual processor survey changed recently to exclude firms that only freeze or wholesale seafood products. This has the effect of removing many scallop processors from the survey.

Figure 4. Time Series of Landing Ranks for the 25 Large Ports Used in the Panel and Cross-Sectional Models. While the Largest Ports are Relatively Stable, the Rank of the Smaller Ports Display a Good Amount of Variability
A Long-Run Growth Perspective
Scallop biomass and landings have increased substantially in the past two decades; we first examine the relative importance of natural advantages and returns-to-scale on long-run growth of scallop landings in ports in the Northeast US. In standard empirical growth models, growth rates in output are inversely related to initial values (Barro and Sala-I-Martin Reference Barro and Sala-I-Martin1992, Mankiw, Romer, and Weil Reference Mankiw, Romer and Weil1992); that is, in the long run, economies with relatively small initial capital stocks will grow more quickly than large economics because the marginal productivity for additional capital in these economies is greater. Applied to a model of scallop landings at the port level, the standard economic growth model regresses the natural log of growth rate (g i, annualized over N years) on initial conditions (Landings i0) and other explanatory variables (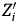 $ Z^{\prime}_i $):
$ Z^{\prime}_i $):
 $$g_i = \displaystyle{1 \over N}\lpar \ln Landings_{iT}-\ln Landings_{i0}\rpar = \alpha + \beta _1\ln Landings_{i0} + \beta _k{Z}^{\prime}_i + \varepsilon _i.$$
$$g_i = \displaystyle{1 \over N}\lpar \ln Landings_{iT}-\ln Landings_{i0}\rpar = \alpha + \beta _1\ln Landings_{i0} + \beta _k{Z}^{\prime}_i + \varepsilon _i.$$Adding 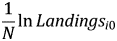 $ \displaystyle{1 \over N}\ln Landings_{i0} $ to both sides of equation 2 and collecting terms produces the equivalent specification:
$ \displaystyle{1 \over N}\ln Landings_{i0} $ to both sides of equation 2 and collecting terms produces the equivalent specification:
 $$\displaystyle{1 \over N}\ln Landings_{iT} = \alpha + \lpar \beta _1 + \displaystyle{1 \over N}\rpar \ln Landings_{i0} + \beta _k{Z}^{\prime}_i + \varepsilon _i.$$
$$\displaystyle{1 \over N}\ln Landings_{iT} = \alpha + \lpar \beta _1 + \displaystyle{1 \over N}\rpar \ln Landings_{i0} + \beta _k{Z}^{\prime}_i + \varepsilon _i.$$ In extensions of the standard growth model, 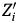 $ Z^{\prime}_i $ typically includes variables that control for stocks of human capital, physical capital, natural capital, institutions, and geography. After controlling for these common steady-state determinants across regions, the β1 coefficient is expected to be negative, and large (or small) absolute values indicate fast (or slow) economic convergence.
$ Z^{\prime}_i $ typically includes variables that control for stocks of human capital, physical capital, natural capital, institutions, and geography. After controlling for these common steady-state determinants across regions, the β1 coefficient is expected to be negative, and large (or small) absolute values indicate fast (or slow) economic convergence.
We aggregate our core dataset into three time periods: initial (t = 0:1996 − 1999), intermediate (t = M:2000 − 2011), and terminal (t = T:2012 − 2015). Initial and terminal conditions are constructed as averages during t = 0 and t = T periods, respectively. This aggregation smooths out year-to-year fluctuations, providing a more general representation of long-run fishery conditions and outcomes during these periods. We focus on the 105 ports that had landings during the four-year period from 2012–2015. Landings in these 105 ports represent just over 99 percent of total landings over the entire study period. An additional 28 ports had landings in the initial period (1996–1999), but not the terminal period.
Two initial conditions, initial scallop landings (Landings i0) and initial nonscallop landings value (Other i0), are constructed that proxy for returns to scale and returns to scope respectivelyFootnote 9. We also construct two long-run natural advantage proxies derived from the available biomass index during the intermediate time period:
 $${AVG}_i = \displaystyle{1 \over N}\sum\limits_{t\in M} \ln Available_{it}$$
$${AVG}_i = \displaystyle{1 \over N}\sum\limits_{t\in M} \ln Available_{it}$$ $${S}{D}_i = \sqrt {\displaystyle{{\sum\limits_{t\in M} {{\lpar \ln Available_{it}-\overline{InAvailable}_{it}} \rpar }^2}} \over {\lpar N-1\rpar }}} $$
$${S}{D}_i = \sqrt {\displaystyle{{\sum\limits_{t\in M} {{\lpar \ln Available_{it}-\overline{InAvailable}_{it}} \rpar }^2}} \over {\lpar N-1\rpar }}} $$The average (AVG i) and standard deviation (SD i) of the Availability Index are both included in our long-run growth models to control for port accessibility to natural capital. It is reasonable to think that increases in Availability would lead to increases in landings; over time, good (poor) access to biomass could induce firms to relocate into (out) of a port. We suspect that variability (SD i) leads to a lower growth rate. We also include controls for activity in other fisheries to explain differences in landings growth across ports. These may work through agglomeration economies if, for example, larger ports are able to attract landings from smaller ports by exploiting economies of scale and thick input markets to offer services at lower costs. Similarly, decreases in fishing activity could result in fewer full-service ports and encourage consolidation of a fishery into a small set of core ports as mobile fishing firms (or fishing rights) migrate away from the periphery.
Equation 2 is typically estimated on large cross-sectional units (countries) using aggregate measure of economic activity for which zeros for initial or terminal conditions do not occur. Because we are estimating a model on small cross-sectional units (ports) on a disaggregate measure of economic activity (scallop fishing), we occasionally observe zeros for either initial or terminal conditions. The methodology outlined in Battese (Reference Battese1997) allows us to include observations that have values of zero for the initial condition variables in our sample and obtain unbiased parameter estimates. We construct the variable 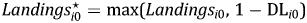 $ Landings_{i0}^{\star} = {\rm max}\lpar Landings_{i0}\comma \; 1-{\rm DL}_{i0}\rpar $, where DLi0 = 1 if Landings i0 > 0 and zero otherwiseFootnote 10. This process is repeated for Other i0 and our estimating equation is:
$ Landings_{i0}^{\star} = {\rm max}\lpar Landings_{i0}\comma \; 1-{\rm DL}_{i0}\rpar $, where DLi0 = 1 if Landings i0 > 0 and zero otherwiseFootnote 10. This process is repeated for Other i0 and our estimating equation is:
 $$\eqalign{& \displaystyle{1 \over N}\ln \lpar Landings_{iT}\rpar = \alpha + \lpar \beta _1 + \displaystyle{1 \over N}\rpar \ln \lpar Landings_{i0}^{\rm \star} \rpar \cr & \quad \quad + \beta _2\ln \lpar Other_{i0}^{\rm \star} \rpar + \beta _3AVG_i + \beta _4SD_i + \beta _5{\rm D}{\rm L}_{i0} + \beta _6{\rm D}{\rm O}_{i0} + {\rm \varepsilon}} $$
$$\eqalign{& \displaystyle{1 \over N}\ln \lpar Landings_{iT}\rpar = \alpha + \lpar \beta _1 + \displaystyle{1 \over N}\rpar \ln \lpar Landings_{i0}^{\rm \star} \rpar \cr & \quad \quad + \beta _2\ln \lpar Other_{i0}^{\rm \star} \rpar + \beta _3AVG_i + \beta _4SD_i + \beta _5{\rm D}{\rm L}_{i0} + \beta _6{\rm D}{\rm O}_{i0} + {\rm \varepsilon}} $$This specification allows us to construct an estimation sample (Estimation Sample 1) that includes ports that had no scallop or other landings in the initial period; however, the 28 ports with no landings in the terminal period still drop out of the sampleFootnote 11.
To examine the robustness of results obtained from specifications that omit these ports, we estimate our growth equations on a second, aggregate estimation sample that aggregates the 28 ports that did not have landings in the terminal period with their nearest neighbors (Estimation Sample 2); summary statistics for both samples are contained in Table 1. Biomass data is recalculated as the annual average across ports for each multiport observation in this aggregated data set.
Table 1. Summary statistics of the variables used in the growth model. Averages with standard deviations below in parenthesis

We extend the specifications defined by equation 6 by interacting the natural advantage variables (AVGi and SDi) with the initial conditions to allow for the effects of natural advantages to vary with initial sizes. For the specifications with interactions, we also de-mean the interaction terms by subtracting the mean values of AVG, SD, Landings, and Other prior to interacting. This facilitate comparisons to the non-interacted model. As part of our specification search, we initially included a market access variable; this did not improve model fit, and we therefore omit these resultsFootnote 12. We also provide estimates based on equation 2 in the appendix using both the aggregate and non-aggregate samples, but note that these estimation samples do not include ports that have values of zero for Landings0 or Other0.
Table 2 display the results of our long-run growth regressions. Columns (1) and (3) show estimates based on the nonaggregated sample of ports; equivalent specifications that use the aggregated sample are shown in columns (2) and (4), respectively. Model fit is reasonably good—R 2 ranges from 0.50 to 0.60 across the four models and when significant, coefficient estimates remain fairly robust. Information criteria approaches for model selection prefers models (1) and (3). However, a joint F-test of the interaction coefficients suggested that the interaction terms belong in the model. Because omitting the explanatory variables can lead to biased estimated coefficients if they truly belong, we focus our interpretation on the results in column (3) of Table 2Footnote 13. The Landings0 coefficient estimated from equation 6 is difficult to interpret: we therefore report the original structural β1 coefficientFootnote 14. The conditional (implied) main effect of initial scallop landings on growth is negative. We interpret this as evidence that, at average levels of the two variables characterizing biomass, ports less engaged in the scallop fishery grew at a faster rate than the larger ones (convergence). This finding is broadly consistent with the finding of increase geographic dispersion by Lee et al. (Reference Lee, Speir, Carr-Harris and Benjamin2017). We also interpret the positive coefficient on non-scallop values as evidence of aggregation economies and returns to scope: ports that were initially active in other fisheries may have been poised to attract scalloping activity.
Table 2. Long-run cross-sectional landings growth models, OLS estimates of Equation 6

The Re-transformed landings coefficient (β1) is reported. All models include a constant term. Nonaggregated port sample used in columns (1) and (3); aggregated port sample used in columns (2) and (4) (see text for details). Robust standard errors in parenthesis. *, **, *** represent significance at the 10 percent, 5 percent, and 1 percent level, respectively.
The positive and statistically significant coefficient on AVG is evidence that better access to biomass leads to higher scallop landings for average-sized ports. However, given the interaction between AVG and the two initial conditions variables, we can examine how this effect varies across the full range of initial port sizes by computing the derivative of growth with respect to AVG at observed values of Landings0 and Other0. The top panels of Figure 5 provide a graphical illustration of these results based on the estimated coefficients from our preferred specification. Values of Landings0 and Other0 are sorted along the horizontal axis in the top left and right panels, respectively. At each specified covariate value of Landings0 and Other0, we estimate the variance of the derivative of growth with respect to AVG using the Delta method; implied derivatives of growth that are statistically significant at the 10 percent level are plotted in black.

Figure 5. Growth Effects Implied by Cross-Section Estimation
The top left panel of Figure 5 shows that the effect of AVG is positive and relatively stable across a range of values for initial scallop landings. The slope of the line fit to these derivatives shows that returns to growth from AVG are slightly amplified for larger scallop ports, which suggests that larger scallop ports are more capable of attracting landings when biomass conditions are favorable than smaller scallop ports. The top right panel of Figure 5 also reveals a positive growth effect from AVG for a majority of ports across the range of initial no-scallop landed revenue. This plot shows a clearly defined relationship between returns to growth from AVG and initial port size, and growth effects that increase in magnitude with initial non-scallop landed revenues. Comparing the results of both plots in the top panel of Figure 5 leads to a somewhat counterintuitive conclusion: the effect of AVG on landings growth is more sensitive to ports' initial engagement in other fisheries than it is to ports' initial engagement in the scallop fishery, but both plots highlight the fact that large ports (those highly engaged in the scallop fishery or other fisheries) are better able to capitalize on long-run biomass conditions. This result can be viewed as evidence in favor of an agglomerative theory of landings activity. However, further research is needed to disentangle the underlying mechanisms that may be driving these results.
The bottom panels of Figure 5 provide equivalent illustrations of the implied growth effect from biomass variability as measured by SD. The bottom left panel shows that among ports with few or no scallop landings during the initial period, this effect is positive and statistically significant. This result suggests that variability in the availability of biomass induces landings growth in ports that might otherwise remain disengaged from the fishery. At values of initial scallop landings around the sample mean, the effect of biomass variability on growth is ambiguous. However, the value of initial scallop landings just above the sample mean is a threshold at which the implied growth effect from SD (when significant) turns negative. Although these above average-sized scallop ports are shown to be most adversely affected by variability in biomass, it is likely that these negative effects are partially offset by gains stemming from returns to scale in other fisheries. The bottom right panel of Figure 5 shows that the implied growth effects from SD are mixed across the range of values of non-scallop landed revenue. For some ports that have values of Other0 above the sample average, these effects are insignificantly or negatively related to landings growth. For others, the implied growth effects from SD are positive and of substantial magnitude. The positive and statistically significant coefficient on the interaction between SD and Other0 seems driven only by a handful of these above-average sized ports for which SD is revealed to be a positive determinant of growth.
Exploring Heterogeneity in Port-level Dynamics
The growth regressions examined long-run links between environmental conditions and the location of scalloping activity. This section further explores the relationship between port level scallop landings and biomass for the twenty-five largest ports. We hypothesize that, in equilibrium, landings in a port at time t are a function of available biomass:
 $$landings_t = c + b\,Available_t + e_t$$
$$landings_t = c + b\,Available_t + e_t$$Directly estimating equation 7 by least squares is complicated by the fact that both biomass availability and landings may be nonstationary, causing a spurious relationship between the two. It is also reasonable to think that the equilibrium relationship between landings and biomass never holds, because other factors of production adjust slowly. Therefore, we model this as an autoregressive distributed lag model (ARDL) of order (p,n):
 $$landings_t = c + \sum\limits_{i = 1}^p {a_i}\,landings_{t-i} + \sum\limits_{i = 0}^n {b_i}\,Available_{t-i} + e_t\comma \; $$
$$landings_t = c + \sum\limits_{i = 1}^p {a_i}\,landings_{t-i} + \sum\limits_{i = 0}^n {b_i}\,Available_{t-i} + e_t\comma \; $$which can be written as an error correction model (ECM):
 $$\eqalign{\Delta landings_t =& \gamma + \alpha _1landings_{t-1} + \sum\limits_{i = 1}^p {\theta _i} \Delta landings_{t-i} \cr&+ \sum\limits_{i = 0}^n {\beta _i} \Delta Available_{t-i} + e_t\comma \; $$
$$\eqalign{\Delta landings_t =& \gamma + \alpha _1landings_{t-1} + \sum\limits_{i = 1}^p {\theta _i} \Delta landings_{t-i} \cr&+ \sum\limits_{i = 0}^n {\beta _i} \Delta Available_{t-i} + e_t\comma \; $$where Δ is the first-differences operator (Pesaran, Shin, and Smith Reference Pesaran, Shin and Smith2000). The γ parameter must also be restricted (γ = c/α 1) for these equations 8 and 9 to be equivalent. The available biomass index will be zero if there is no fishable biomass in the water. Since biomass is essential to producing landings, it seems reasonable to specify a model in which c = γ = 0. However, we also specify a model that contains a (restricted) constant as a robustness check. We further assume that biomass availability can affect landings, but that lagged values of biomass availability do not directly affect landings. This seems reasonable: fishermen use current – not past – values and locations of biomass, to catch fish. The ARDL and ECM formulations therefore simplify to:
 $${landing}{\rm s}_t = c + \sum\limits_{i = 1}^p {a_i} {landing}{\rm s}_{t-i} + b_0{Availabl}{\rm e}_t + e_t$$
$${landing}{\rm s}_t = c + \sum\limits_{i = 1}^p {a_i} {landing}{\rm s}_{t-i} + b_0{Availabl}{\rm e}_t + e_t$$ $$\eqalign { \Delta {landing}{\rm s}_t &= {\rm \gamma} + {\alpha} _1\,{landing}{\rm s}_{t-1} + \sum\limits_{i = 1}^p {{\theta} _i} \Delta {landing}{\rm s}_{t-i} \cr&\quad + {\beta} _0\Delta {Availabl}{\rm e}_t + e_t$$
$$\eqalign { \Delta {landing}{\rm s}_t &= {\rm \gamma} + {\alpha} _1\,{landing}{\rm s}_{t-1} + \sum\limits_{i = 1}^p {{\theta} _i} \Delta {landing}{\rm s}_{t-i} \cr&\quad + {\beta} _0\Delta {Availabl}{\rm e}_t + e_t$$ The adjustment speed is 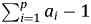 $ \sum\nolimits_{i = 1}^p {a_i-1} $ in equation (10) and equivalently α1 in equation (11). Values near 0 imply slow convergence to the long run equilibrium while values near −1 imply very fast convergence. θ i captures short-run effects while β 0 is the instantaneous marginal effect of available biomass on landings. The long-run marginal effect of available biomass on landings can be computed by dividing β 0 by (1 − α1).
$ \sum\nolimits_{i = 1}^p {a_i-1} $ in equation (10) and equivalently α1 in equation (11). Values near 0 imply slow convergence to the long run equilibrium while values near −1 imply very fast convergence. θ i captures short-run effects while β 0 is the instantaneous marginal effect of available biomass on landings. The long-run marginal effect of available biomass on landings can be computed by dividing β 0 by (1 − α1).
Our estimation strategy has three parts. First, we employ the bounds testing procedure developed by Pesaran, Shin, and Smith (Reference Pesaran, Shin and Smith2001) to test for the existence of a long-run relationship between landings and biomass. To construct the Pesaran, Shin, and Smith (Reference Pesaran, Shin and Smith2001) test, we estimate equation 11. We used the Bayesian Information Criteria (BIC) for model selection, estimating models where p = 1 or 2. The bounds test involves forming an F-statistic for the null hypothesis that α 1, γ, and β 0 are jointly zero. High values of the F-statistic are evidence of a long-run relationship and low values are evidence of no long-run relationship, regardless of whether landings or biomass are integrated of order zero or one. However, there is an inconclusive zone; if the test statistic falls within this zone, then knowledge of the order of integration is required for inference. The critical values are also based on the number of independent variables in equation 11 and the way the constant or time-trends are modeled; we use the critical values reported in Narayan (Reference Narayan2005) and Pesaran, Shin, and Smith (Reference Pesaran, Shin and Smith2001).Footnote 15 As part of this stage, we check that the residuals from equation 11 are not autocorrelated. We will also examine stationarity of landings and biomass using an augmented Dickey-Fuller test (Dickey and Fuller Reference Dickey and Fuller1979).
Second, for ports that have a long-run relationship, we estimate model 10 using OLS and calculate an elasticity of landings with respect to biomass for both models based on port-level averages. Third, for ports that do not have a long-run relationship, we difference equation 7 to produce:
 $$\Delta landings_t = b\Delta Available_t + e_t$$
$$\Delta landings_t = b\Delta Available_t + e_t$$and estimate a short-run model, using OLS. The b terms are the short-run effects of changes in available biomass.
The two versions of the bounds test find that five ports have a long-run relationship between biomass availability and landings: Gloucester, MA, Point Pleasant, NJ, Barnegat, NJ, Wildwood, NJ, and Cape May, NJ (Table 3). There is also some weaker evidence that Harwich, MA and Southampton, NY also have a long-run relationship; however, this relies on the “no constant” specification being correct. Chatham, MA, New London, CT, Chincoteague, VA, and Southampton, NY all have test statistics that fall in the inconclusive region that requires knowledge of the integration order of the landings and biomass. With the exception of landings in Southampton, NY, the augmented Dickey-Fuller stationarity tests fail to reject the null of nonstationarity at the 5 percent confidence level for any port's landings or biomass indexFootnote 16. Because the Dickey-Fuller tests indicate nonstationary landings and available biomass for these ports, we interpret the F-statistics for these ports as evidence of no long-run relationship. The bounds test implies no long-run relationship for the other ports, a somewhat surprising finding, given the results of the growth models.
Table 3. Bounds Tests for a long-run relationship between biomass and landings based on an Error Correction Model with up to two lags; lag length selected using BIC and a Breusch-Godfrey test for no serial correlation of residuals

Tests for stationarity of landings and available biomass at the port level. Ports arranged north to south. The Case I model for Hampton was mis-specified using one or two lags of the dependent variable.
a Evidence of a long-run relationship between biomass and landings at the 5 percent confidence level.
b Evidence of stationarity at a 5 percent confidence level.
We estimate equation 11 without a constant for the five ports with a long-run relationship, plus the two ports for which the evidence is mixed (Table 4)Footnote 17. Model fit is reasonable: R 2 range from 0.42 to 0.58. The BIC values are used for model selection within ports but are not directly comparable across ports. Ports without a ΔLandingst−1 coefficient in Table 4 correspond to an AR(1) model. The coefficient on landingst−1 is the speed of adjustment: this varies nearly threefold from the port that adjusts fastest (Gloucester) to the port that adjusts the slowest (Barnegat). For all seven ports, increases in available biomass result in higher landings, as expected.
Table 4. Model results, diagnostics, and derived long-run effects of available biomass on scallop landings for an ARDL with at most 2 lags of the dependent variable and no constant term

Standard errors for the long-run effects are computed using the delta method; elasticities are computed at average values. Ports are arranged north to south.
The fit of the short-run models ranges from nil to reasonable; R2 range from less than 0.01 to 0.49 (Table 5). One of the more interesting findings is that the simple short-run model explains little variability in landings for the New England ports, but performs reasonably well for the Mid-Atlantic ports. The statistically significant coefficients on the available biomass index are positive.
Table 5. Model results and diagnostics for a short-run model (Equation 12)

Standard errors for four ports (Barnegat, Atlantic City, Chincoteague, and Hampton) computed using Newey-West standard errors due to autocorrelation of the residuals. The “Rank” column reports rank of that port in terms of scallop value over the time series and is presented for ease of interpretation.
a Breusch-Godfrey statistic suggest autocorrelated residuals at the 10 percent significance level.
Chincoteague, VA is the exception; this port had relatively high scallop landings in the early 2000s, 2010, and 2014–2015 (Figure 4) when the biomass index was relatively low (Figure 3a). Conversely, scallop landings were relatively low from the mid 2000s through 2012 (except 2010), when biomass was high. This strong inverse relationship is quite surprising. One explanation could be that fishing vessels that use this port have a particularly strong preference to continue operating from that port while vessels using the other ports are more footloose.
Discussion and Conclusions
In this research, we have characterized some aspects of the economic geography of the sea scallop fishery along the northeast coast of the United States. Our results indicate that factors related to economic geography and port size affect the long-run dynamics of landings patterns. Exploratory analysis indicated that smaller ports have more variable rank orders than larger ports (Figure 4) a finding consistent with Watson and Johnson (Reference Watson and Johnson2012). Our models of long run growth in port-level landings in Section 4 provide evidence that ports with lower initial scallop landings have grown the fastest, a result consistent with theories of dispersion from regional science and results in Lee et al. (Reference Lee, Speir, Carr-Harris and Benjamin2017). Furthermore, like Portman, Jin, and Thunberg (Reference Portman, Jin and Thunberg2009, Reference Portman, Jin and Thunberg2011), we also find evidence that port-level natural advantages influence long-run changes in landings, although this effect exhibits considerable heterogeneity across ports. Access to high average biomass values is correlated with higher rates of landings growth in larger ports. Higher variability in available biomass tends to result in lower rates of long run growth in scallop landings for larger ports.
While our long run growth models in Section 4 suggest that access to biomass is an important determinant of landings in ports our port-level time series models in Section 5 show a high degree of heterogeneity in the response of scallop landings to biomass. The time-series models show relationships between landings and biomass in both large and small ports; we find long-run, positive relationships between biomass and landings for the 2, 5, 7, 14, 20, 21, and 24 ranked ports. Short-run models have reasonable explanatory power for the 2nd-8th ranked ports. Like the long-run models, the magnitudes of marginal effects of biomass on landings vary widely across the ports. The time-series models do not have explanatory power for the moderate-sized ports located in New York, Connecticut, and Massachusetts, nor the largest port (New Bedford).
We suspect that heterogeneity in the natural productivity of parts of the ocean and the distances from ports to those areas (natural advantages) combined with fisheries management to induce a spatial component to the port-level response to changes in biomass availability. Ports in the southern portion of our study area (say, Point Pleasant, NJ and further south) tended to have positive short-term relationships between biomass and landings. The biomass areas in the Mid-Atlantic Bight (from LI south, see Figure 1) are characterized by generally lower abundance with spikes in abundance in some years (see Figure 2). These southern areas may be characterized by temporary high and low levels of scalloping activity when the nearby fishing grounds are not good alternatives or are closed to fishing entirely. In contrast, ports further north, adjacent to biomass areas off the coast of New England, tend to have access to better fishing grounds that are characterized by both higher abundances and less intrayear variation. When a port's closest option is unavailable, it may still be viable to land scallops even if the fishing grounds are more distant. Fishery managers, concerned with the short-run effects of regulations on these southern communities, might consider regulations that smooth the interyear variability in access to biomass or regulations that lower the cost of access for these southern ports. For example, managers might open a southern area to fishing before scallops are optimally sized or construct a no-cost transit corridor (New England Fishery Management Council 2015). These policies would have consequences for economic efficiency.
Returns to scope (from the growth equations) suggest that ports with other fishing activities could be well-positioned to attract new fishing activity when stock conditions are favorable. While we have only characterized one direction by which spillovers might operate, it is possible that scallop fishing could crowd out or attract other fishing activity. While beyond the scope of this study, examining the extent to which these effects spill out to the broader marine economy would provide tremendous insight into the way fisheries management can affect regional economies. For example, if dock or other space is at a premium, then fisheries that are highly profitable may crowd out the less profitable ones. Alternatively, traditional spillover mechanisms, like thick inputs markets may counteract this effect. Local governments and other stakeholders have been concerned about the preservation of shoreside infrastructure, working waterfronts, and social capital in the fishing industry (Breen and Rigby Reference Breen and Rigby1985, Georgianna et al. Reference Georgianna, Avila, Bethoney, Cass, Challingsworth and Hogan2014, Ounanian Reference Ounanian2015). There are theoretical reasons to believe the locations of at least some fishery support infrastructure (including seafood processors, repair facilities, and supply shops) both affects and is affected by landings location (and therefore biomass availability). For example, processors may choose to locate in locations with high levels of landings to take advantage of economies of scale and reduce the transport costs of raw materials. Alternatively, they may locate in closer to final demand if transport costs of raw materials are low. Further research characterizing the mechanisms by which fishing activities affect associated industries and communities is essential for fisheries managers to fully understand the effects of management on fishing communities.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/age.2018.23
Acknowledgements
The authors are grateful for comments from participants at the 2016 IIFET and NAREA conferences and from the anonymous reviewers. The findings and conclusions in the manuscript are those of the authors and do not reflect the position of the Department of Commerce or the National Oceanographic and Atmospheric Administration. As usual, all remaining errors are our own.













 Open access
Open access