Introduction
Afro-Palearctic long-distance migrant birds are declining faster than sedentary birds with which they share breeding habitats (Vickery et al. Reference Vickery, Ewing, Smith, Pain, Bairlein, Skorpilova and Gregory2014). One possible explanation for this differential decline may be the particular conditions that affect migrants in their wintering grounds and/or in their passage areas (Sanderson et al. Reference Sanderson, Donald, Pain, Burfield and van Bommel2006). In long-distance migratory birds, mortality rates can be considerably higher during migration than during sedentary periods (Sillett and Holmes Reference Sillett and Holmes2002, Terraube et al. Reference Terraube, Mougeot, Cornulier, Verma, Gavrilov and Arroyo2012, Klaassen et al. Reference Klaassen, Hake, Strandberg, Koks, Trierweiler, Exo, Bairlein and Alerstam2014) and reproductive success can be influenced by the quality of habitats located thousands of kilometres away in their non-breeding grounds (e.g. Norris et al. Reference Norris, Marra, Kyser, Sherry and Ratcliffe2004). Moreover, due to their dependency on disjointed areas that they are likely subjected to diverse conservation policies, migrant birds usually found less protection during their annual cycle compared with resident species (Runge et al. Reference Runge, Watson, Butchart, Hanson, Possingham and Fuller2015). It is therefore critical to improve our knowledge of the ecology, distribution and habitat use of migratory birds outside their breeding season in order to elaborate effective holistic conservation programs (Martin et al. Reference Martin, Chades, Arcese, Marra, Possingham and Norris2007).
In recent decades, breeding populations of the European Roller Coracias garrulus (hereafter, Roller) have suffered among the most drastic declines for Afro-Palearctic migrant farmland birds wintering in open savannas (Sanderson et al. Reference Sanderson, Donald, Pain, Burfield and van Bommel2006). The annual cycle of Rollers is distributed across their breeding grounds (April–August), non-breeding grounds (November–February) and migratory pathways (August–November for autumn migration and February–April for spring migration) (Rodríguez-Ruiz et al. Reference Rodríguez-Ruiz, De la Puente, Parejo, Valera, Calero-Torralbo, Reyes-Gonzalez, Zajková, Bermejo and Avilés2014, Finch et al. Reference Finch, Saunders, Avilés, Bermejo, Catry, De la Puente, Emmenegger, Mardega, Mayet, Parejo, Racinskis, Rodríguez-Ruiz, Sackl, Schwartz, Tiefenbach, Valera, Hewson, Franco and Butler2015). In several countries in Europe, covering most of the global breeding range, the species has become extinct or is declining (Burfield and Van Bommel Reference Burfield and Van Bommel2004, Kovacs et al. Reference Kovacs, Barov, Orhun and Gallo-Orsi2008). The rate of global population decline of Rollers has lessened but not stopped in the last decade, and its conservation status changed recently from ‘Near Threatened’ to ‘Least Concern’ (Burfield and Van Bommel Reference Burfield and Van Bommel2004, Kovacs et al. Reference Kovacs, Barov, Orhun and Gallo-Orsi2008, BirdLife International 2015). The reasons for the decline and fragmentation of breeding populations include the loss of suitable habitats, agricultural intensification and misuse of pesticides in breeding areas (Avilés and Parejo Reference Avilés and Parejo2004, Kovacs et al. Reference Kovacs, Barov, Orhun and Gallo-Orsi2008). However, there is still an important lack of information on the threats faced by Rollers during migration and on their non-breeding grounds, impeding us from quantifying the relative importance of the different threats year-round.
Recently, the use of geolocators and satellite telemetry has allowed locating the non-breeding grounds of Rollers from the westernmost part of the Eurasian breeding range (France, Portugal and Spain). Rollers from these breeding populations spend the non-breeding season in an area located in southern Angola, northern and central Namibia and northern Botswana (Emmenegger et al. Reference Emmenegger, Mayet, Duriez and Hahn2014, Catry et al. Reference Catry, Catry, Granadeiro, Franco and Moreira2014, Rodriguez-Ruiz et al. Reference Rodríguez-Ruiz, De la Puente, Parejo, Valera, Calero-Torralbo, Reyes-Gonzalez, Zajková, Bermejo and Avilés2014). Rollers breeding in Bulgaria, Cyprus, Latvia and Montenegro, however, spend the non-breeding season farther east, between Botswana, Zambia, Zimbabwe, Tanzania and Kenya (Finch et al. Reference Finch, Saunders, Avilés, Bermejo, Catry, De la Puente, Emmenegger, Mardega, Mayet, Parejo, Racinskis, Rodríguez-Ruiz, Sackl, Schwartz, Tiefenbach, Valera, Hewson, Franco and Butler2015). Bird atlases extend the Roller non-breeding distribution towards the south, with highest densities in the north-east of South Africa, and exclude the most arid areas in the south-west of Africa (Keith et al. Reference Keith, Urban and Fry1988, Herremans et al. Reference Herremans, Brown, Borello and Herremans-Tonnoeyr1993). The most south-western part is most likely occupied by the subspecies C. g. semenowi, that breeds from Iraq to north-western China (Cramp Reference Cramp1998). Tracking studies from across Europe reveal that Rollers from different breeding populations use different but overlapping non-breeding areas (“moderate connectivity”; Finch et al. Reference Finch, Saunders, Avilés, Bermejo, Catry, De la Puente, Emmenegger, Mardega, Mayet, Parejo, Racinskis, Rodríguez-Ruiz, Sackl, Schwartz, Tiefenbach, Valera, Hewson, Franco and Butler2015). In such a scenario, it is critical to characterise the habitat use and environmental variables that determine species distribution during the non-breeding season in order to identify potential threats and deliver effective conservation actions for this species. It is also important to assess to what extent the most suitable non-breeding areas are included within the current network of protected areas (PAs). Land cover change is the major threat affecting loss of biodiversity (Sala et al. Reference Sala, Chapin, Armesto, Berlow, Bloomfield, Dirzo, Huber-Sandwald, Huenneke, Jackson, Kinzig, Leemans, Lodge, Mooney, Oesterheld, Poff, Sykes, Walker, Walker and Wall2000) and PAs have been proved as an effective measure to reduce loss of natural land cover (Beresford et al. Reference Beresford, Eshiamwata, Donald, Balmford, Bertzky, Brink, Fishpool, Mayaux, Phalan, Simonetti and Buchanan2013). Assessing the degree of coverage of protected areas on the wintering grounds of Rollers would help to identify potential gaps in protection and key areas for its conservation.
Here we aim to contribute to the conservation of Rollers by identifying key areas and land cover used by Rollers during winter from two different perspectives: from population-specific to the overall species. For that purpose, we develop two different ecological niche models based on climatic, topographic and habitat variables to identify suitable wintering areas for Rollers in southern Africa: first, using satellite telemetry data obtained from individuals marked during breeding in Spain to determine the extent of potential areas used by the Spanish breeding population (hereafter Spanish population) and, second, using online open-access databases on Roller occurrences as input for the model, to identify the requirements and distribution range for the overall species, including Rollers from different breeding origins across Europe (hereafter overall population). Finally, we use the model outputs to evaluate to what extent the most suitable non-breeding areas and land cover types are included within the current network of PAs in Africa.
Methods
Occurrence data from satellite tracking
We used satellite-tracking occurrence data from six adult birds (four males, one female and one unsexed individual) that were trapped during the 2012 and 2013 breeding seasons in Spain at six different locations (provinces of Badajoz, Ciudad Real, Girona, Granada, Lleida and Madrid). Each tagged bird provided only one wintering period. Birds were equipped with 5 g solar-powered PTT-100 satellite transmitters (Microwave Telemetry Inc., Columbia, MD, USA) that were fitted with a backpack Teflon harness (Garcelon Reference Garcelon1985). The device to body-weight ratio was less than 5% (Rodríguez-Ruiz et al. Reference Rodríguez-Ruiz, De la Puente, Parejo, Valera, Calero-Torralbo, Reyes-Gonzalez, Zajková, Bermejo and Avilés2014).
Satellite transmitters were programmed with an 8h ON⁄ 15h OFF duty cycle. We used only high-quality locations, classified according to their accuracy as LC 3 (up to 250 m), 2 (up to 500 m) or 1 (up to 1.5 km) (Argos 2007) to describe non-breeding areas. The start of the wintering period was defined as the day when the bird stopped displacement for more than 10 days. The end of the wintering period was when the bird moved more than 20 km in a day with a bearing between NW and NE (see Rodríguez-Ruiz et al. Reference Rodríguez-Ruiz, De la Puente, Parejo, Valera, Calero-Torralbo, Reyes-Gonzalez, Zajková, Bermejo and Avilés2014 for more details). We filtered locations to avoid pseudo-replication issues associated with repeated locations of a given individual in the same grid cell; for this, we overlapped a grid of 15 arc-min cell size (roughly 25×25 km) and selected one random location per cell, obtaining a total of 33 locations (1–9 locations per bird).
Occurrence data from online databases
In order to describe the non-breeding areas used by the overall population of Rollers in southern Africa (irrespective of their breeding origin), we used occurrence data available from the Southern African Bird Atlas Project (SABAP1; Harrison et al. Reference Harrison, Allan, Underhill, Herremans, Tree, Parker and Brown1997). Although there is currently an ongoing second phase of the SABAP atlas, to date the SABAP1 has a more complete coverage of our study area and was selected for this reason. We complemented this dataset with available records from the open source database GBIF (Global Biodiversity Information Facility; http://www.gbif.org/). In the GBIF database, we discarded records from bird collections and kept only data 1) with reported location accuracy; 2) from 2000 to 2014 in order to describe recent Roller distribution, 3) collected between 1 December and 28 February to ensure that observations belong to individuals residing on their non-breeding area (Rodriguez-Ruiz et al. Reference Rodríguez-Ruiz, De la Puente, Parejo, Valera, Calero-Torralbo, Reyes-Gonzalez, Zajková, Bermejo and Avilés2014, Table 1) and not to individuals recorded while on passage that may not be actually staying in the area (Lee and Barnard Reference Lee and Barnard2017); and 4) occurring within the southern African range (latitude: 15° to 36° S; longitude: 11° to 34° E). Records of Rollers wintering north of 15°S are anecdotal, and restricted to very few locations, so sampling cannot be considered representative for modelling (see model requirements below). In total, we built a dataset consisting of 1,167 occurrences obtained from these sources, which we used to model the potential suitable distribution of the overall population in southern Africa.
Table 1. Variables used in the ecological niche models.
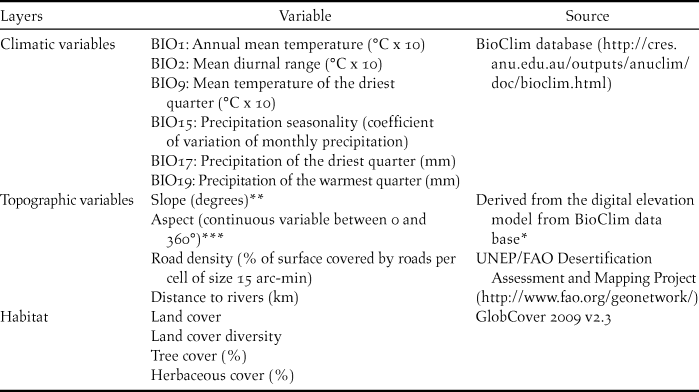
* Calculated with QGIS (QGIS 2015)
** Calculated according to the eight surrounding pixels for every grid cell
*** Pixels with a slope less than 2° were considered as flat (no aspect)
Environmental data
We used 14 environmental variables to model the ecological niche of the species. We obtained climatic variables from the WorldClim database (Hijmans et al. Reference Hijmans, Cameron, Parra, Jones and Jarvis2005), which includes 19 bioclimatic predictor layers summarising annual trends, seasonality and extremes in temperature and precipitation. As many of these variables were correlated, we extracted the values from all the 19 bioclimatic predictors corresponding to our occurrence records and selected six independent climate variables for analyses (using regression trees and variance inflation factor analyses to avoid multi-collinearity among selected variables). Besides climatic, topographic and habitat variables, distance to rivers and road density were included for their a priori biological relevance, as these are known to influence habitat preferences of Rollers during breeding (Cramp Reference Cramp1998, Rodriguez et al. Reference Rodríguez, Avilés and Parejo2011). A detailed list of variables is in Table 1.
Spatial resolution
All environmental variables were rescaled by overlaying them with a grid of 15 arc-min cell size (approximately 25×25 km) including almost all sub-equatorial Africa (see coordinates range considered above). This scale was chosen to fit the resolution of the SABAP1 census method that included most of the locations of wintering Rollers in southern Africa. The resampling of the data to adjust the scale was performed using the nearest-neighbour method for the categorical variable land cover and the bilinear interpolation method for the continuous variables. For the road density variable, we used the number of roads per cell at the given resolution.
Ecological niche models
We modelled the ecological niche of Rollers in southern Africa using the MaxEnt software (version 3.3.3k; Phillips et al. Reference Phillips, Anderson and Schapire2006) implemented in the R package dismo. MaxEnt uses presence-only data to estimate the probability distribution of a species and a maximum entropy modelling approach (Phillips et al. Reference Phillips, Anderson and Schapire2006; Elith et al. Reference Elith, Phillips, Hastie, Dudik, Chee and Yates2011). It also requires a set of background points where species occurrence was not measured to contrast against presences, from which it predicts the environmental conditions occupied by the species in proportion to their availability in the landscape (Elith et al. Reference Elith, Phillips, Hastie, Dudik, Chee and Yates2011). MaxEnt is also reported to have the best performance when dealing with small sample sizes (Hernández et al. Reference Hernández, Graham, Master and Albert2006). This method works under two key assumptions: a) sampling is either random or representative throughout the landscape, and b) detection probability is constant across sites (Yackulic et al. Reference Yackulic, Chandler, Zipkin, Royle, Nichols, Grant and Veran2013). By using satellite-tracking data these two assumptions are met, as tracked birds represent a random sample of the breeding population. These assumptions are also fulfilled by data from online databases. Indeed, the SABAP1 had a constant effort along the area considered in this study and Rollers are conspicuous birds that almost invariably perch in exposed places (i.e. trees and/or pylons) close to bush-cleared grassland in southern Africa (Herremans et al. Reference Herremans, Brown, Borello and Herremans-Tonnoeyr1993), thus making their detection an easy task.
We created a set of 1,000 points randomly distributed over the considered extent of southern Africa using the function randomPoints included in dismo package in R, checking for non-overlap with presences (i.e. grey points in Figure 1). Occurrence and background datasets were separated into five groups. We randomly selected four of these groups for model fitting (80% of data for training) and one group for testing the resultant model (20% of the data), alternatively changing the training and testing groups for each replication. The models were run using logistic output format, with 500 iterations set for model convergence. Variable importance was measured using the jackknife method. Models performances were evaluated using the area under the receiver-operating curve (AUC) (Fielding and Bell Reference Fielding and Bell1997), for which values above 0.75 are usually considered as of good performance and values above 0.90 are considered very good. Regulation multiplier (β) was previously selected by exploring a pool of models using a range of β values from 1 to 10 in steps of 0.5. β was chosen attending to AUC, AICc and smooth response curves.
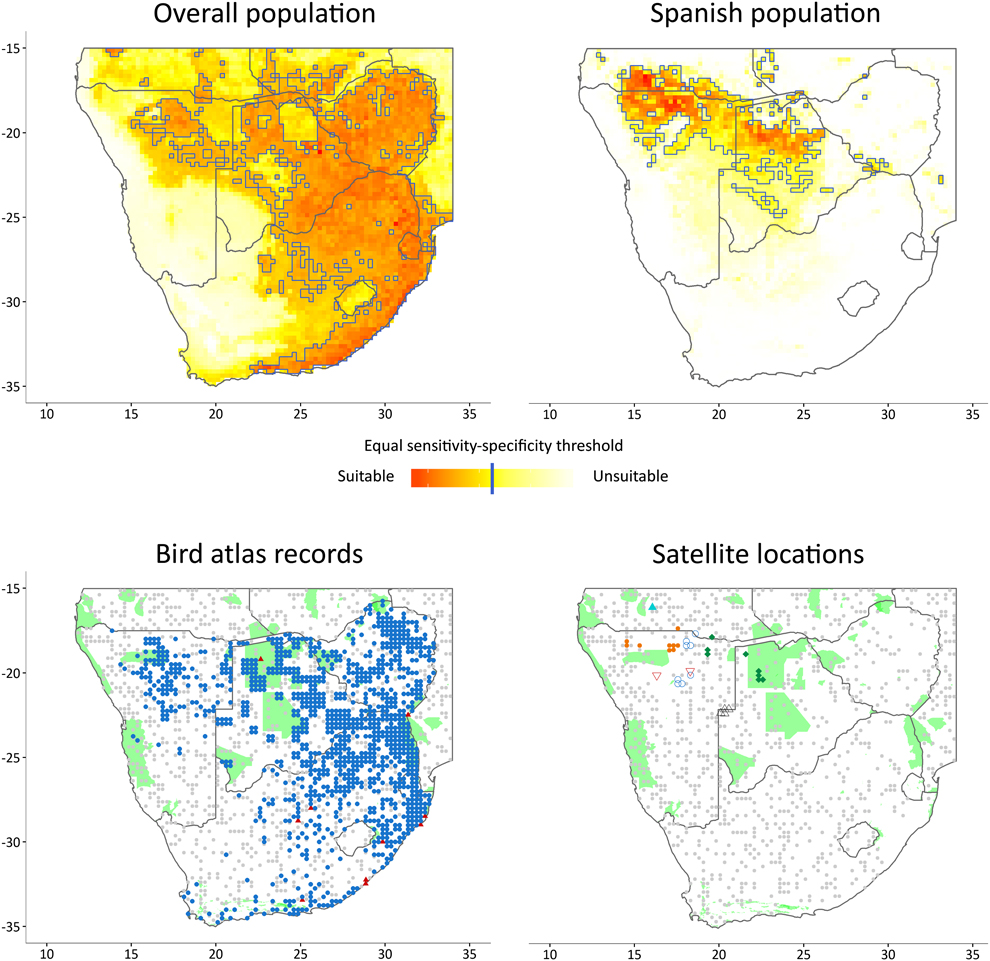
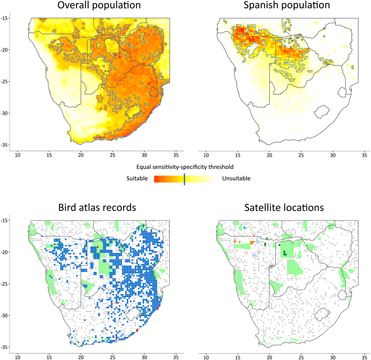
Figure 1. Results of the ecological niche models for the overall population (upper-left) and for the Spanish population (upper-right) in Southern Africa. Map at bottom-left shows the records from the SABAP (blue dots) and GBIF (red triangles) database and their associated background points (grey dots). Map at bottom-right shows the locations of the satellite-tracked Rollers (individuals in different shape and colour) and their associated background points (grey dots). The existing protected areas with international recognition are shown in green in the two maps at bottom (UNEP-WCMC 2010).
MaxEnt produces a map showing the relative suitability of species occurrence, giving a value for every grid cell that ranges from 0 to 1. We used the equal sensitivity-specificity threshold (ESS) as a criterion of suitability: regions of the map with values above this threshold were considered as suitable for the species (Bean et al. Reference Bean, Stafford and Brashares2012, Cao et al. Reference Cao, DeWalt, Robinson, Tweddale, Hinz and Pessino2013). We evaluated the relative contribution of variables to the model and how they affected the model predictions attending to the response curves given by MaxEnt.
Land cover use
Firstly, we use the 33 filtered satellite locations of tracked Spanish Rollers to identify which land cover types are actually used during the wintering period and whether there are individual preferences. We also used the generated maps to estimate the proportions of land cover types used by the overall population and by the Spanish population. We did so by superposing the suitable areas for the species (occurrence probability > ESS) onto the GlobCover 2009 v2.3 land cover map using R.
Protected areas
We assessed the extent to which suitable areas for the species overlapped with extant PAs, using nationally recognized PAs listed in the 2010 World Database on Protected Areas (IUCN and UNEP-WCMC 2010). We only considered PAs with IUCN classifications, Ramsar Convention Sites, World Heritage Sites, UNESCO-MAB Biosphere Reserves and sites within other international conventions. A total of 95 PAs covered 11.08% of the total study area (Figure 1).
Results
Ecological niche models
The identified suitable wintering area for the Spanish population was relatively narrow and located mainly in the south of Angola, north of Namibia and central Botswana, with scattered small locations in Zambia, Zimbabwe and Mozambique (Figure 1). The climatic variables that contributed most to the ecological niche model (> 5%) were mean annual temperature and precipitation in the driest quarter (11.68 and 8.69 %, respectively; Table 2). Mean annual temperature was positively correlated to probability of occurrence, with a turning point at 23°C. Precipitation in the driest quarter was inversely correlated until 40 mm, when the curve stabilizes (Figure 2). The non-climatic variables that contributed most to the model were land cover, slope and tree cover (34.33%, 24.27% and 15.67%, respectively; Table 2). The land cover type with highest probability of occurrence was the savanna. Probability of occurrence decreased with increasing slope; when the slope was greater than 2% the probability of occurrence was almost zero. Probability of occurrence increased with tree cover until 5%, but then decreased above this threshold value (Figure 2). The discriminatory power of the model for the Spanish population was very good, as assessed by the area under the curve (AUC = 0.927).
Table 2. Model parameters and contribution (%) of environmental variables to the ecological niche models. Two separate models were developed to identify suitable areas for the overall population and for the Spanish population of Rollers. Most important variables in each model are shown in bold (i.e. Variable importance > 5%, in terms of contribution to the model).
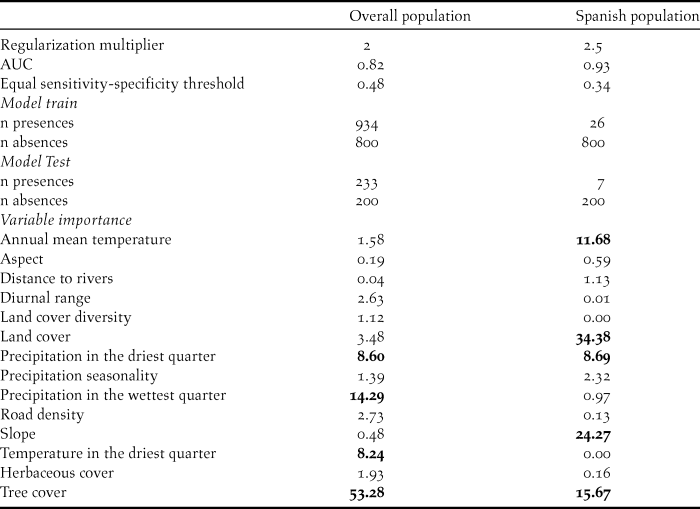
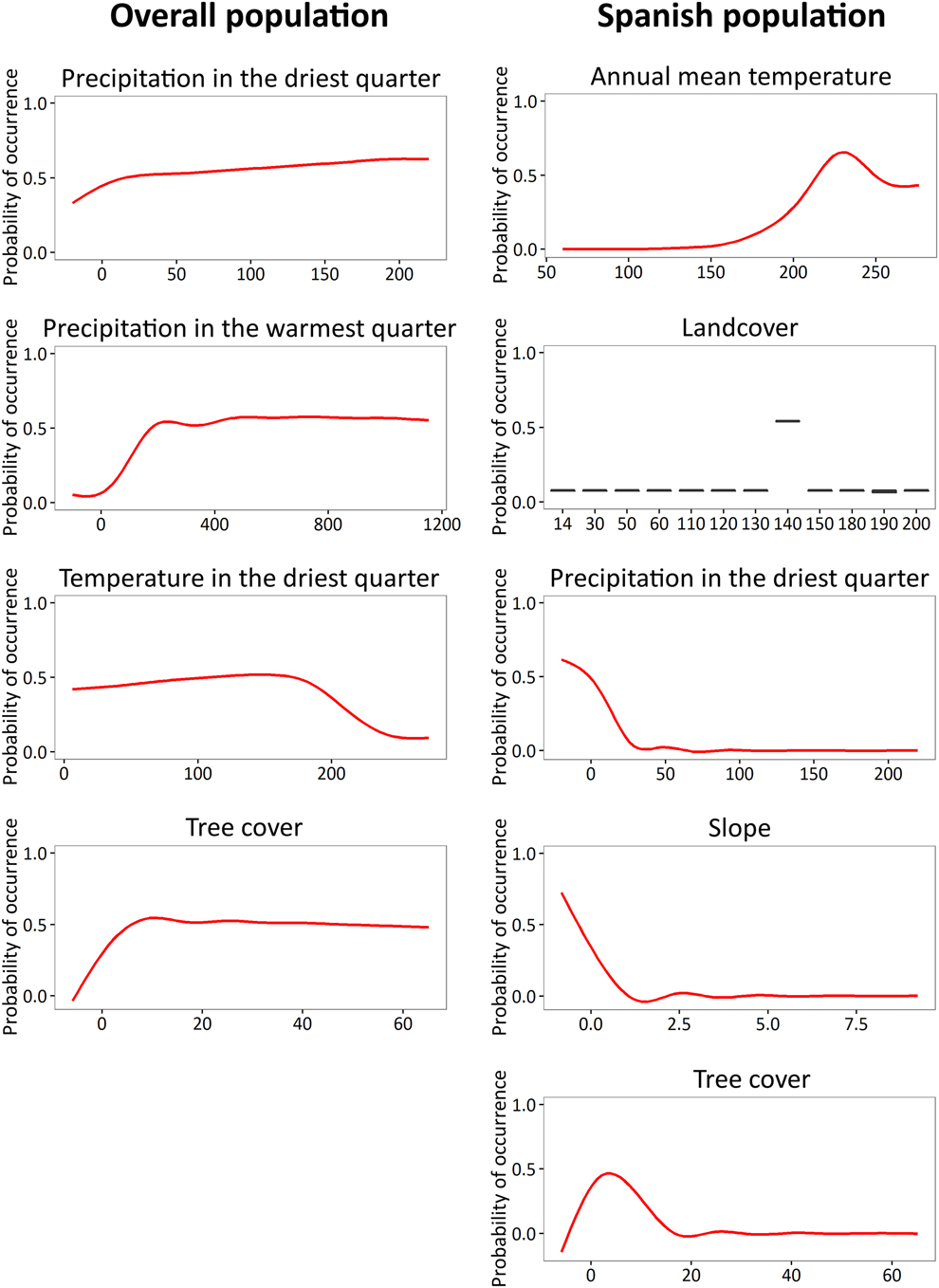
Figure 2. Partial response curves representing the relation between probability of occurrence of the Spanish population (left) and the overall population (right) and the five most important factors contributing to the models. Units are given in Table 2.
The identified suitable wintering areas for the overall population (irrespective of breeding origin) consisted mainly of the eastern part of southern Africa, Zimbabwe, eastern Botswana and the north-east and east coast of South Africa, as well as isolated patches in the north of Namibia (Figure 1). The most important variables contributing to this model were tree cover, temperature in the warmest quarter, precipitation in the driest quarter and temperature in the driest quarter (53.28%, 14.29%, 8.6% and 8.24%, respectively; Table 2). Tree cover was initially positively correlated to probability of occurrence, reaching a turning point around 10% of coverage, when the curve stabilises. The three climatic variables were positively correlated with occurrence probability, but precipitation in the warmest quarter and temperature in the driest quarter reached turning points at 200 mm and 18° C, respectively (Figure 2). The model for the overall population also performed well (AUC = 0.818).
Land cover use
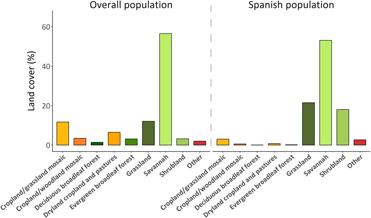
Three land cover types were selected by the tracked Rollers: grassland, savanna and shrubland, yet we found individual differences among them (χ210 = 18.74, P = 0.04). Five individuals used mainly grassland and savanna, whereas one roller used only shrubland. For models, considering both the Spanish population and the overall population, we found that the savanna was the primarily used land cover (53.10% and 56.55%, respectively), followed by grasslands (21.52% and 12.12%). Shrublands also appeared to be important for the Spanish population (18.06%), whereas mosaics of croplands/grasslands and dry croplands were important for the overall population (11.72% and 6.45%, respectively). All these land cover types represented more than 85% of the total predicted suitable areas (Figure 3).
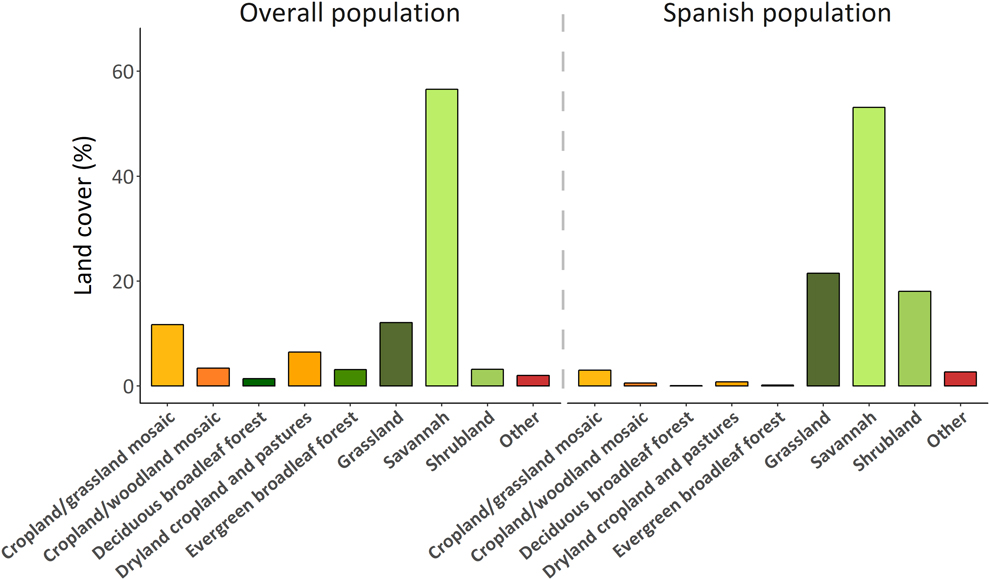
Figure 3. Proportion of land cover areas included in the optimal distributions predicted by MaxEnt for the overall population and for the Spanish population. Greenish colours represent non-managed land cover types, yellowish colours represent man-managed land cover types.
Protected areas
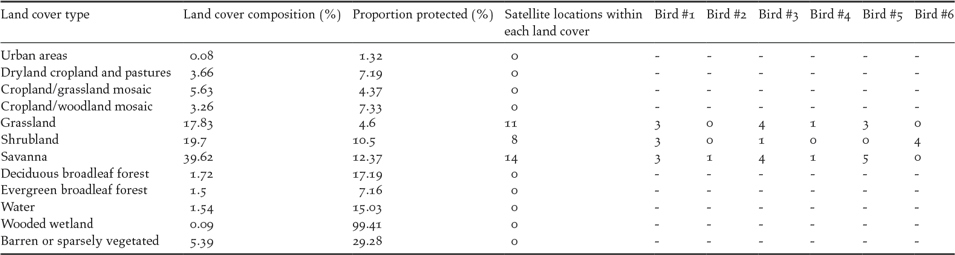
Only one tracked bird actually used PAs during its wintering period (four out of eight locations inside PAs), yet birds often wintered in places close to PAs: 18 out of 33 locations were within 100 km of a PA boundary, and the distance between satellite locations and the nearest PA was 66.7 km (range 0–223.9). For the Spanish population, 18.37% of the identified suitable area was included within PAs (26 of 95 PAs; see Table 4 for details of the most relevant PAs). For the overall population, however, only 9.58% of the predicted suitable area was included within PAs (66 of 95 PAs).
Savanna, grassland and shrubland represent 77% of the total study area, but only 10% of this area was covered by PAs. The proportion of these land covers that was protected compared with the proportion that was outside PAs did not differ from that of other land cover types, such as pastures or cropland (χ211 = 14.724, P = 0.195), unlike wetlands, which were almost entirely protected (Table 3).
Table 3. Land cover composition in study area, protected areas coverage of each land cover type and number of satellite locations from tracked birds that falls within each land cover type.
Discussion
Suitable non-breeding areas for Rollers
To our knowledge, this is the first study combining two different perspectives (i.e. satellite-tracking and bird atlases) to estimate and identify suitable areas and potential threats to an endangered migratory bird species. The suitable non-breeding areas for the Spanish population were located in the north-western part of southern Africa, whereas the overall population occupied a wider region, from central southern Africa to the east and from north to the south along the eastern coast of Africa. We found little overlap between the two predicted suitable areas. However, comparisons must be made carefully, because input data for each model came from different sources and hence likely had different probabilities of detection. The under-representation of the species in the north-western region in the model for the overall population may be a consequence of a differential sampling effort in the SABAP1 in these regions, or of biases due to the limitations of satellite data from only six individuals. A higher sample of tracked individuals, perhaps including Rollers breeding in populations close to the ones sampled here, and a higher participation in the censuses for the most remote areas in southern Africa could provide a more unbiased representation of the distributional range of the species during wintering. Nonetheless, these results support the previous patterns found using tracking devices suggesting that birds from different breeding populations spend their winter in different but overlapping areas (Finch et al. Reference Finch, Saunders, Avilés, Bermejo, Catry, De la Puente, Emmenegger, Mardega, Mayet, Parejo, Racinskis, Rodríguez-Ruiz, Sackl, Schwartz, Tiefenbach, Valera, Hewson, Franco and Butler2015). From our data, we infer that birds breeding in south-western populations migrate to the northwestern part of the total non-breeding range area for the species. Therefore, birds from other breeding populations (i.e. Central or Eastern Europe) are probably those spending the non-breeding season farther east (see Figure 1– map of the overall population). This idea is supported by a recent study of the migration of Rollers breeding in Bulgaria, Cyprus, Latvia and Montenegro, showing that these birds winter in southern Africa farther east than Rollers breeding in Spain (Finch et al. Reference Finch, Saunders, Avilés, Bermejo, Catry, De la Puente, Emmenegger, Mardega, Mayet, Parejo, Racinskis, Rodríguez-Ruiz, Sackl, Schwartz, Tiefenbach, Valera, Hewson, Franco and Butler2015).
The ecological niche models indicate that the non-breeding distribution of Rollers is strongly influenced by climatic variables. Both precipitation and temperature had an important weight when predicting the distribution of Rollers. Therefore, changes in the general patterns of these climatic variables might affect the extent of suitable areas for the species through indirect changes in habitat or in prey abundance (Huntley et al. Reference Huntley, Collingham, Green, Hilton, Rahbek and Willis2006, Reference Huntley, Collingham, Willis and Green2008, Chen et al. Reference Chen, Hill, Ohlemuller, Roy and Thomas2011). However, this pattern is not consistent when considering the Spanish population, for which climatic variables appeared less relevant. This may be due to the less diverse climatic conditions in a more limited band of latitude, which makes other variables, such as land cover, more relevant. Precipitation in the driest quarter was an important predictor in the models of the Spanish population and the overall population of Rollers, although the tolerance to this variable (i.e. response curve) differed. Indeed, the Spanish population showed a narrower range of tolerable conditions (between 0 and 25 mm) than the overall population (between 0 and 200 mm; Figure 2), perhaps because the response curve for the overall population describes the summed tolerance of each separate breeding population. Two alternative explanations for these different thresholds in the wintering areas are possible: 1) individuals track niches with conditions and resources more similar to those of their breeding grounds, driven by population-specific adaptations (Price and Gross Reference Price, Gross, Greenberg and Marra2005, Monahan and Tingley Reference Monahan and Tingley2012); or 2) populations distribution in the wintering quarters reflects the longitudinal distribution in their breeding areas following a parallel migration (Finch et al. Reference Finch, Saunders, Avilés, Bermejo, Catry, De la Puente, Emmenegger, Mardega, Mayet, Parejo, Racinskis, Rodríguez-Ruiz, Sackl, Schwartz, Tiefenbach, Valera, Hewson, Franco and Butler2015), making the climatic conditions found in Africa a collateral consequence and not a real tolerance range.
Besides climatic variables, tree cover appears as an important factor that influences the ecological niche and distribution of both the overall population and the Spanish population. Suitability is higher when tree cover is low for the Spanish population, while the tree cover threshold appears to be higher for the overall population (Figure 2). Rollers usually use open landscapes with sparse trees in their breeding grounds (Avilés and Costillo Reference Avilés and Costillo1998) and this preference also seems to occur during the non-breeding season, at least for the Spanish population. In the case of the Spanish population, slope and land cover are the most important variables determining the suitable area. Spanish Rollers seem to be restricted to flat areas and savanna. Even though Rollers are known to choose breeding territories next to watercourses (Cramp Reference Cramp1998), which usually present slopes, these conditions do not necessarily apply to non-breeding areas, where Rollers may find flat areas with scattered trees as a suitable landscape due to the availability of perches and a better line of sight to forage.
Roller land cover use in southern Africa
During the non-breeding season (December–February), Spanish Rollers used land cover types characterised by a high proportion of natural vegetation (savanna, grassland and shrubland, but the predicted suitable areas for the overall population overlapped with a higher proportion of anthropogenically managed land cover types (Figure 3). This pattern may be explained by the relative abundance of land cover types within each particular range. In the eastern part of southern Africa, a high proportion of savanna, grassland and shrubland has been progressively converted into crops due to high human population density (Lambin et al. Reference Lambin, Turner, Geist, Agbola, Angelsen, Bruce, Coomes, Dirzo, Fischer, Folke, George, Homewood, Imbernon, Leemans, Li, Moran, Mortimore, Ramakrishnan, Richards, Skanes, Steffen, Stone, Svedin, Veldkamp, Vogel and Xu2001, Linard et al. Reference Linard, Gilbert, Snow, Noor and Tatem2012). In these low-intensive farming areas, the replacement of natural savanna leads to large increases in the availability of prey species for insectivorous birds (e.g. grasshoppers; Hulme Reference Hulme2007). However, this also means more insect pests for crops and a greater use of pesticides in cultivated areas to combat these, which may have an indirect negative effect on insectivorous birds in general, and Rollers in particular (Everts Reference Everts1990, Mullie and Keith Reference Mullie and Keith1993, Avilés and Parejo Reference Avilés and Parejo2004, Newton Reference Newton2004). Organochlorine pesticides (DDT and DDE) have been recently detected in avian predators from southern Africa, indicating that these compounds still represent a risk, either through an illegal use outside malaria areas or their long persistence in the environment (García-Heras et al. Reference García-Heras, Simmons, Arroyo, Camarero, Mateo and Mougeot2018).
Overlap with conservation areas
Savanna, grassland and shrubland are proportionally less protected than wetlands, but are also very important to migratory birds like Rollers. In fact, this is not the first study highlighting the need of increasing the protection of crucial habitats for migratory birds of semi-arid areas (Limiñana et al. Reference Limiñana, Arroyo, Terraube, McGrady and Mougeot2015). Our results show that the current network of PAs covers only a tenth of the non-breeding areas identified as most suitable for the overall population of Rollers in southern Africa. The modelled optimal distribution of the Spanish population, however, benefited from a relatively greater degree of protection, with almost 20% of most suitable areas being currently included within PAs. Compared with other studies in Africa conducted on species with similar requirements, there is still room for improvement in protection coverage (Limiñana et al. Reference Limiñana, Arroyo, Terraube, McGrady and Mougeot2015), although Rollers find a better scenario than other species (Limiñana et al. Reference Limiñana, Soutullo, Arroyo and Urios2012). Interestingly, Rollers also used areas close to PA boundaries. The reasons for this should be elucidated, but this observation suggests that a small expansion of the PAs could be a benefit for this species. The relatively high occurrence of Rollers outside PAs has several mutually non-exclusive explanations: 1) current PAs are mainly designed to protect charismatic megafauna, and, therefore, have an insufficient coverage of suitable habitats for long-distance migrant birds; 2) unprotected areas are attracting Rollers due to their high availability of perches for hunting (e.g. power lines along roads). A previous study has found that availability of perch sites is a factor favouring breeding territory choice for Spanish Rollers (Avilés and Costillo Reference Avilés and Costillo1998). PAs in Africa are in areas with low human population density and, therefore, have a reduced network of roads and associated power lines and may provide Rollers with a low number of perches (see above); 3) unprotected areas might be attracting Rollers because competition with resident insectivores could be lower than inside PAs (Herremans Reference Herremans1998); or 4) because the higher agricultural intensification in unprotected areas leads to an increase of plant material that facilitates the accidental introduction of foreign insect arthropod pests (Abate et al. 2000) which may constitute new feeding opportunities for Rollers.
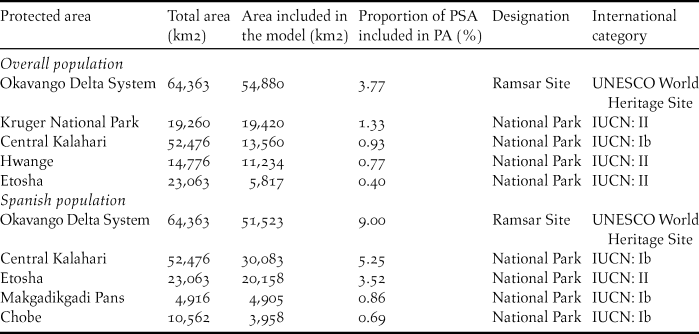
Between 74% and 82% (Spanish population and overall population, respectively) of the estimated suitable area for Rollers which is under coverage of PAs is categorised as National Park or Ramsar site. The Okavango Delta System in Botswana appears to be one PA that contains larger proportions of predicted suitable area for both the Spanish population and the overall populations of Rollers (Table 4). This PA is listed as a Ramsar and UNESCO World Heritage Site, however, its conservation status is threatened by climate change and an increasing pressure to satisfy the demand for water and grazing lands of a growing surrounding population (BirdLife International 2018a). Focusing on Rollers, threats that may compromise the suitability of this area are overgrazing, clear-cutting and the use of insecticides that could reduce prey availability (Thieme et al. Reference Thieme, Abell, Burgess, Lehner, Dinerstein and Olson2005). The Kruger National Park in north-eastern South Africa, Kalahari National Park in central Botswana and Etosha National Park in north-western Namibia, also protect large suitable areas for Rollers, although only Kalahari enjoys a favourable conservation status (BirdLife International 2018b; 2018c; 2018d). Between 18% and 26% of the suitable area for Rollers under protection are categorised as minor parks (e.g. nature reserves and game reserves) and their conservation status and their contribution to the welfare of Rollers in them should be assessed carefully.
Table 4. List of the five most important protected areas contributing to the predicted suitable areas (PSA) for the overall population and for the Spanish population of Rollers.
Summing up, PAs in southern Africa cover, on average, a low proportion of the suitable habitat for threatened species (Beresford et al. Reference Beresford, Buchanan, Donald, Butchart, Fishpool and Rondinini2011, Runge et al. Reference Runge, Watson, Butchart, Hanson, Possingham and Fuller2015). This seems to be also the case for the European Roller, until very recently considered as ‘Near Threatened’ by IUCN and currently considered as ‘Vulnerable’ in Spain. Given the extensive area used by the overall population of Rollers during wintering, conservation measures directed to manage the landscape on a broad scale may be preferable to the investing the limited economical resources dedicated to conservation into extending the existing PA network. For instance, developing agricultural policies that reduce intensification and protect the grasslands would be beneficial. We also point out that different breeding populations may face different conservation threats in their respective non-breeding areas and that a conservation plan focusing on the most suitable areas for the species in general may not be sufficient to protect all breeding populations. We highlight the importance of analysing the particular requirements of each population to elaborate an integrated and effective conservation plan for the species as a whole, which is more relevant, if possible, when there is some migratory connectivity, as in the case of the Roller (Finch et al. Reference Finch, Saunders, Avilés, Bermejo, Catry, De la Puente, Emmenegger, Mardega, Mayet, Parejo, Racinskis, Rodríguez-Ruiz, Sackl, Schwartz, Tiefenbach, Valera, Hewson, Franco and Butler2015). In this sense, attending to the wide predicted wintering area, international cooperation is essential to make a joined effort in fulfilling the conservation needs of this species, as well as those of other sub-Saharan migrants. Further research is also needed for locating key stopover areas for different Roller populations and assessing their protection status during this critical stage of their life cycle.
Acknowledgements
We thank Dan Chamberlain, Tom Finch, Graeme Buchanan and two anonymous reviewers for their comments that helped in shaping this manuscript. DP and JMA were funded by the Spanish Ministry of Education and Science/FEDER through the projects CGL2008-00718 and by the Government of Extremadura through the project TA13002 to DP. All the information about tagged Rollers in Spain has been obtained in the framework of the ‘Migra’ program (www.migraciondeaves.org/en/) developed by SEO/BirdLife, coordinated by Juan Carlos del Moral, and financed by Iberdrola Foundation. The Extremadura Regional Government provided information on one individual. Manuel Aguilera, Jaume Bonfil, Gerard Bota, Pedro Bustamante, Jesús Fernández, José Antonio Fimia, María José Herrero, Jesús Morena, Juan Carlos Perlado, Sergio Romero and Chema Traverso helped during fieldwork. Domingo Rivera, José María Abad and Toribio Álvarez (Junta de Extremadura) made fieldwork and downloaded all the information from the individual from Extremadura.