



Understanding Pleistocene subsistence strategies is critical to developing models of mobility and to answering questions about how humans settled North America. Current models of human subsistence during the late Pleistocene (>11,700 cal BP) in North America are predominantly derived from and biased toward faunal data. Although Paleoindian food economies undoubtedly included plants, the paucity of archaeobotanical assemblages from well-dated contexts limits our view of dietary diversity and foraging choices. Recent studies, however, have recovered fragile dietary remains from Pleistocene-aged contexts, reinforcing the need for a paleoethnobotanical perspective for understanding variability in Paleoindian lifeways.
Early models of Paleoindian land use centered on evidence from archaeological sites in the Great Plains and Southwest (Haynes Reference Haynes1966; Heizer and Baumhoff Reference Heizer and Baumhoff1970; Martin Reference Martin1973), where extinct megafaunal remains including mammoth (Mammuthus columbi) and bison (Bison antiquus) were associated with fluted-point technology (Figgins Reference Figgins1933; Sellards Reference Sellards1952). Researchers posited an adaptive strategy focusing on specialized big-game hunting and high mobility (Kelly and Todd Reference Kelly and Todd1988; Surovell and Waguespack Reference Surovell, Waguespack and Haynes2009), with some claiming that early Paleoindian groups targeted megafauna to the point of extinction (Martin Reference Martin1973). Others have challenged this interpretation (e.g., Cannon and Meltzer Reference Cannon and Meltzer2004), and rigorous debate continues regarding the importance of large mammals in Paleoindian subsistence (Grayson and Meltzer Reference Grayson and Meltzer2003; Haynes and Hutson Reference Haynes, Hutson, Graf, Ketron and Waters2013; Meltzer Reference Meltzer2020; Waguespack Reference Waguespack, Graf, Ketron and Waters2013).
The role of plants in early Paleoindian lifeways has received significantly less attention. The skew in the archaeological discourse toward hunting over gathering is at least partially due to the dearth of Pleistocene-aged archaeobotanical assemblages, especially in temperate North America where less than 10% of Pleistocene sites with preserved subsistence material have yielded substantial (more than five botanical specimens) and unequivocal dietary plant remains (Figure 1). Our comprehensive evaluation of archaeobotanical data from Pleistocene-aged sites in North America (see Supplemental Text 1) demonstrates that current evidence for Paleoindian plant consumption is scarce—no doubt due to several factors including preservation, sampling, and the relative importance of such resources in people's diets. The record has grown in the last two decades, however, and reliable evidence for dietary plant use is known from five sites in the American Northeast, Southeast, and Great Basin, where archaeobotanical assemblages are of sufficient size and context to confidently interpret dietary use. Botanical data from these sites confirm that some early Paleoindian groups foraged for diverse plant species and suggest that regionally adapted subsistence strategies were in place by at least the Younger Dryas (ca. 12,900–11,700 cal BP).
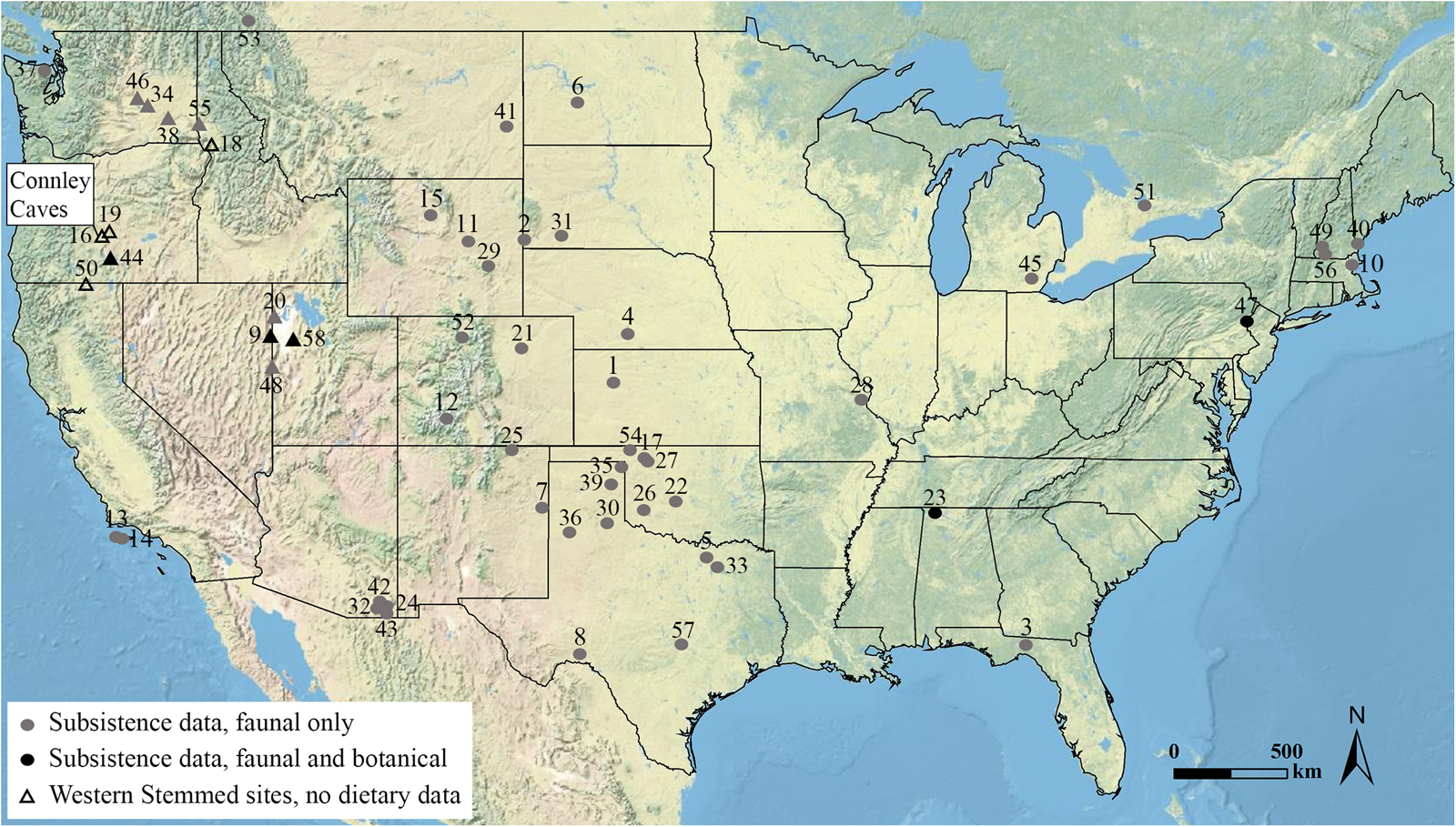
Figure 1. Map displaying location of Connley Caves and Paleoindian sites with subsistence data older than 11,700 cal BP (subsistence associations for faunal data are from Anderson et al. Reference Anderson, Smallwood and Shane Miller2015; Gingerich and Kitchel Reference Gingerich, Kitchel, Smallwood and Jennings2015; Grayson and Meltzer Reference Grayson and Meltzer2015; Haynes and Hutson Reference Haynes, Hutson, Graf, Ketron and Waters2013; Hill Reference Hill2008; Hurst et al. Reference Hurst, Carter and Athfield2010; Kilby et al. Reference Kilby, Farrell and Hamilton2021; Lothrop et al. Reference Lothrop, Lowery, Spiess and Ellis2016; Mackie et al. Reference Mackie, Surovell, O'Brien, Kelly, Pelton, Vance Haynes and Frison2020; botanical data with total counts of less than five seeds were considered unreliable and excluded from the map). Sites with Western Stemmed technology are displayed as triangles, and sites with other tool technologies are displayed as circles. 1: 12 Mile Creek; 2: Agate Basin; 3: Alexon; 4: Allen; 5: Aubrey; 6: Big Black; 7: Blackwater Draw; 8: Bonfire Shelter; 9: Bonneville Estates Rockshelter; 10: Bull Brook; 11: Casper; 12: Cattle Guard; 13: Channel Islands SMI-261; 14: Channel Islands SRI-512; 15: Colby; 16: Connley Caves; 17: Cooper; 18: Cooper's Ferry; 19: Cougar Mountain Cave; 20: Danger Cave; 21: Dent; 22: Domebo; 23: Dust Cave; 24: Escapule; 25: Folsom; 26: Howard Gully; 27: Jake Bluff; 28: Kimmswick; 29: La Prele; 30: Lake Theo-Folsom; 31: Lange-Ferguson; 32: Lehner; 33: Lewisville; 34: Lind Coulee; 35: Lipscomb; 36: Lubbock Lake; 37: Manis; 38: Marmes Rockshelter; 39: Miami; 40: Michaud; 41: Mill Iron; 42: Murray Springs; 43: Naco; 44: Paisley Caves; 45: Pleasant Lake; 46: Sentinel Gap; 47: Shawnee Minisink; 48: Smith Creek Cave; 49: Tenant Swamp; 50: Tule Rockshelter; 51: Udora; 52: Upper Twin Mountain; 53: Wally's Beach; 54: Waugh; 55: Wewukiyepuh; 56: Whipple; 57: Wilson Leonard; 58: Wishbone.
At the Clovis site of Shawnee Minisink, Pennsylvania, Gingerich (Reference Gingerich2011) contends that charred hawthorn fruit and hickory nuts found in hearths provide the most compelling evidence of dietary plant use and attributes those remains to opportunistic foraging near camp (but see Dent Reference Dent, Walker and Driskell2007). At Dust Cave, Alabama, charred botanical remains from Younger Dryas-aged components containing Quad/Beaver Lake and Dalton projectile-point technology suggest people consumed a variety of nuts (hickory [Carya sp.], black walnut [Juglans nigra], acorn [Quercus sp.], hazelnut [Corylus sp.]), berries (blackberry [Rubus sp.], and grape [Vitis sp.]) and possibly seeds (Chenopodium sp.; Hollenbach Reference Hollenbach, Walker and Driskell2007). These two sites provide a view of plant use east of the Rocky Mountains indicating that late Pleistocene foragers procured nuts and fruits. In the American Great Plains, Southwest, Colorado Plateau, and California, clear evidence of plant foods is absent prior to the Early Holocene (Bement et al. Reference Bement, Drass, Cummings and Larrick2020; Gill et al. Reference Gill, Braje, Smith and Erlandson2021; Janetski et al. Reference Janetski, Bodily, Newbold and Yoder2012; Louderback Reference Louderback2014; Louderback and Pavlik Reference Louderback and Pavlik2017; Rosenthal and Fitzgerald Reference Rosenthal, Fitzgerald, Bousman and Vierra2012; Yoder et al. Reference Yoder, Bodily, Hill, Janetski and Newbold2010; see Supplemental Text 1).
Three sites in the Great Basin have reliable evidence of Pleistocene plant consumption. At Paisley Caves, Oregon, charred plants in Pleistocene-aged hearths suggest that people consumed small seeds from grasses (Poaceae), goosefoot (Chenopodium sp.), cheno-ams (Amaranthaceae), tansymustards (Descurainia sp.), phacelia (Phacelia sp.), and borages (Boraginaceae), as well as parenchymous tissue of geophytes (Kennedy Reference Kennedy2018). Younger Dryas-aged coprolites contained amaranth (Amaranthaceae) seeds, rose fruit (Rosaceae), high frequencies of legume (Fabaceae) pollen, and common occurrences of phytoliths, indicating the regular consumption of leafy greens (Blong et al. Reference Blong, Adams, Sanchez, Jenkins, Bull and Shillito2020). At Bonneville Estates Rockshelter, Nevada, hearths dated between ~13,000 and 11,700 cal BP contained charred cactus (Opuntia sp.) parts and small seeds including grasses (Leymus sp., Achnatherum hymenoides, and Sporobolus sp.), goosefoot, sunflower (Asteraceae), bulrush (Schoenoplectus sp. or Bolboschoenus sp.), and mustard (Brassicaceae), all interpreted as economic (Rhode and Louderback Reference Rhode, Louderback, Graf and Schmitt2007). At Wishbone, Utah, charred botanicals in a ~12,300 cal BP hearth indicate economic use of red maid (Calandrinia sp.), pitseed goosefoot (Chenopodium berlandieri), and tobacco (Nicotiana sp.; Duke et al. Reference Duke, Rice, Young and Byerly2018, Reference Duke, Wohlgemuth, Adams, Armstrong-Ingram, Rice and Craig Young2021). All three sites contain Haskett technology (Smith et al. Reference Smith, Duke, Jenkins, Goebel, Davis, O'Grady, Stueber, Pratt and Smith2020), suggesting that early stemmed-point makers had diverse diets including plants.
With the findings from Paisley Caves, Bonneville Estates Rockshelter, and Wishbone, the Great Basin region has taken center stage in the discussion of Paleoindian plant consumption. In this region, people used Western Stemmed Tradition (WST) hunting technology coeval with the Clovis Tradition (Davis et al. Reference Davis, Madsen, Becerra-Valdivia, Higham, Sisson, Skinner and Stueber2019; Jenkins et al. Reference Jenkins, Davis, Stafford, Campos, Hockett, Jones, Cummings, Yost, Connolly and Yohe2012; Shillito et al. Reference Shillito, Whelton, Blong, Jenkins, Connolly and Bull2020), and WST forms continued to be made through the Younger Dryas (Goebel and Keene Reference Goebel, Keene, Janetski and Parezo2014; Rosencrance Reference Rosencrance2019; Smith et al. Reference Smith, Duke, Jenkins, Goebel, Davis, O'Grady, Stueber, Pratt and Smith2020). Despite the growing Paleoindian record, WST sites still lack unequivocal evidence for the hunting of extinct megafauna (Goebel et al. Reference Goebel, Hockett, Adams, Rhode and Graf2011). Instead, current models predict that groups using WST technology hunted some large mammals but overall employed more diverse subsistence strategies than did groups on the Great Plains (Elston et al. Reference Elston, Zeanah and Codding2014; Hockett Reference Hockett, Graf and Schmitt2007; Madsen Reference Madsen2015). The degree of resource diversity and its relationship to settlement patterns, however, are still debated (e.g., Duke and Young Reference Duke, Craig Young, Graf and Schmitt2007).
Recent research at Connley Caves, a multicomponent WST site located in central Oregon, offers a new opportunity to gain a diachronic view of Paleoindian plant use. Here we present archaeobotanical data and 17 new radiocarbon ages from five combustion features spanning the Pleistocene–Holocene Transition (PHT) in Connley Cave 5 (PHT; ca. 13,000–8200 cal BP). Our objectives are to (1) characterize plant use and foraging activities at Connley Caves during the PHT, (2) consider how these data fit into the growing body of archaeobotanical data for the Pleistocene Paleoindian period more broadly, and (3) discuss implications for current settlement-subsistence models.
Connley Caves
Connley Caves are located on the traditional lands of the Klamath, Modoc, and Northern Paiute in the Fort Rock Basin of central Oregon. The Fort Rock Basin is the northwesternmost drainage system in the Great Basin, encompassing an area of ~3,900 km2 bordered by the High Lava Plains (to the north), Cascade foothills (west), and woodland marshes of the Klamath Basin (south). Ridgelines divide the basin into three valleys: Silver Lake, Christmas, and Fort Rock. The site comprises at least eight rockshelters situated in the south-facing slope of Connley Hills overlooking Paulina Marsh in the Silver Lake Valley (Figure 2). Its location at the intersection of two physiographic regions and its position between the valley floor and uplands allow ready access to a variety of habitats—providing a rare look at foraging behavior in an ecotonal setting (Aikens and Jenkins Reference Aikens and Jenkins1994; Prouty Reference Prouty, Aikens and Jenkins1994). The caves are ~45 m above the valley floor (~1,356 m asl) within a woodland belt of western juniper (Juniperus occidentalis) and a sparse understory dominated by sagebrush (Artemisia), rabbitbrush (Chrysothamnus), and grasses such as bluebunch wheatgrass (Pseudoroegneria spicata), Idaho fescue (Festuca idahoensis), and western needlegrass (Achnatherum occidentale; Bureau of Land Management 2000; Franklin and Dyrness Reference Franklin and Dyrness1988). Juniper woodland transitions into a sagebrush steppe community at the base of the Connley Hills, followed by dune formations and alkaline playa that support salt desert shrub communities of greasewood (Sarcobatus vermiculatus), shadscale (Atriplex confertifolia), and budsage (Artemisia spinescens) along the northern margin of Paulina Marsh. Emergent herbs and grass-like plants including bulrushes (e.g., Scirpus, Schoenoplectus), sedges (Carex, Cyperus), and cattail (Typha) are common throughout the marsh. Paulina Marsh's mosaic of wetland patches and meadows is a critical source of water for flora and fauna.
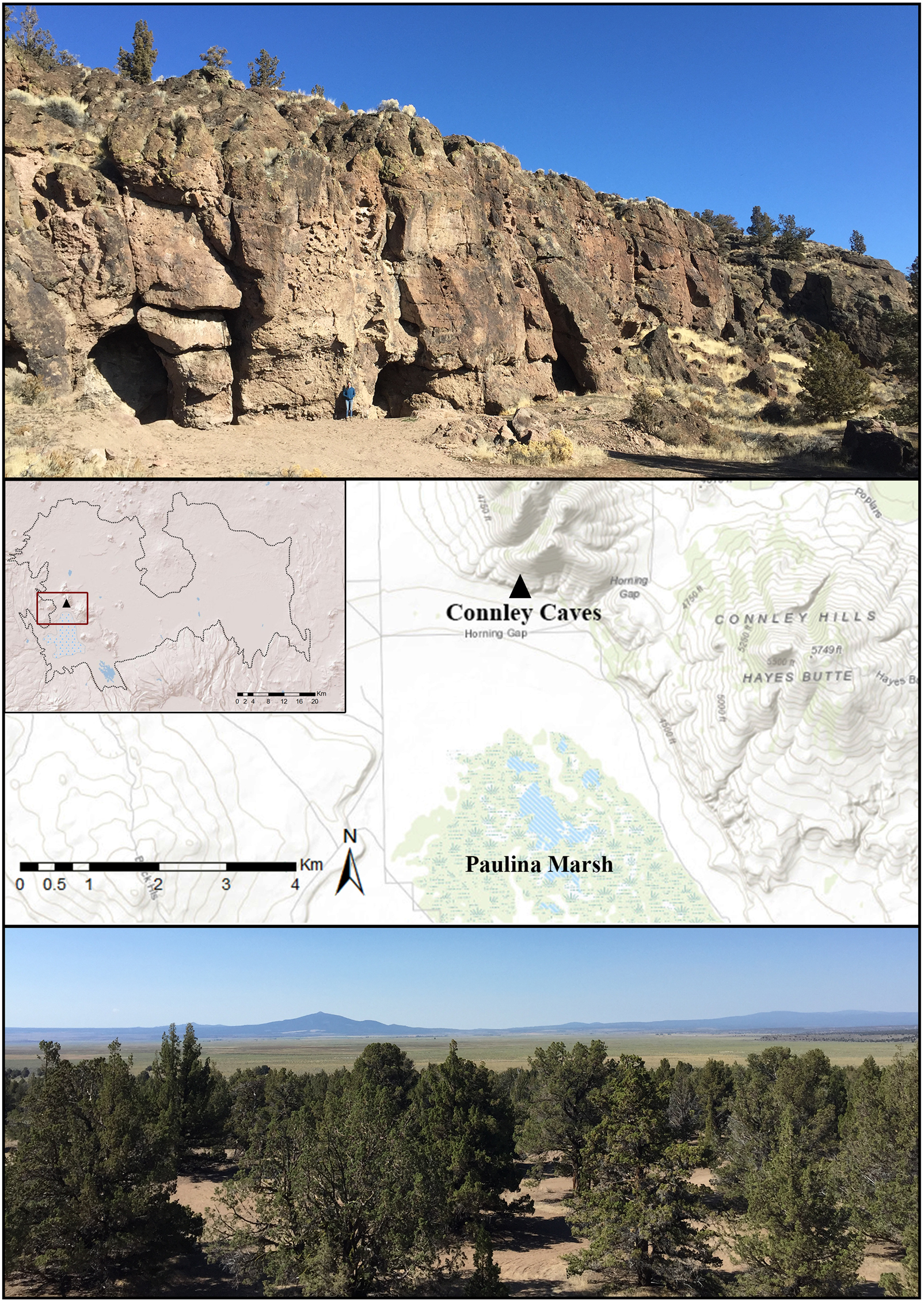
Figure 2. Top: View of Connley Caves looking north–northeast with person in blue standing above the Cave 5 excavation block. Middle: Location of the site within the Fort Rock Basin showing proximity to Paulina Marsh. Bottom: Southward view of Paulina Marsh with Hager Mountain from the top of Connley Caves (photos by Richard Rosencrance). (Color online)
The site gained recognition in the archaeological community after Stephen Bedwell's excavations in the 1960s that uncovered artifacts in purported association with charcoal dating to ~13,000 cal BP (Bedwell Reference Bedwell1970). Connley Caves also played an important role in the development of Bedwell's Western Pluvial Lakes Tradition concept, which proposed that WST groups in the western Great Basin were “tethered” to lacustrine settings. For more than 50 years, scholars have speculated about the cultural chronology of Connley Caves (Bryan Reference Bryan, Harten, Warren and Tuohy1980; Grayson Reference Grayson, Sheets and Grayson1979), with some archaeologists questioning whether they were heavily used prior to the Early Holocene (Goebel et al. Reference Goebel, Hockett, Adams, Rhode and Graf2011:493). The Archaeology Field School of the University of Oregon returned to the site between 2000–2001 and 2014–2021 to refine its chronological, ecological, and technological records (Jenkins et al. Reference Jenkins, Holcomb and McDonough2017; McDonough et al. Reference McDonough, Rosencrance, Holcomb and Jenkins2018). Excavations in Connley Caves 4 and 5 uncovered multiple cultural components spanning the Younger Dryas through the Early Holocene (ca. 11,700–8200 cal BP; Walker et al. Reference Walker, Head, Lowe, Berkelhammer, Björck, Cheng and Cwynar2019). Here we provide the first look at the Paleoindian components recently excavated in Cave 5, which contain extensive WST lithic and osseous tool assemblages, discrete activity areas, and cultural features.
Materials and Methods
This study examines sediment samples collected between 2017 and 2019 in Connley Cave 5, including 11 samples from inside five combustion features, two comparative samples from outside the features, and 17 control samples from an associated sediment column (Figure 3). Combustion features represent unique archaeological contexts that can inform about human behavior at the individual scale, reflecting a single event or repeated use over weeks, months, or years. Such features can take many forms in archaeological sites, and we follow the definitions outlined by Mentzer (Reference Mentzer2014) to differentiate hearths, hearth areas, and combustion areas. Supplemental Table 1 provides additional details about features and sampling methods. To establish chronological control, we submitted piece-plotted charcoal from within each combustion feature to the Pennsylvania State University AMS Radiocarbon Laboratory and Direct AMS for radiocarbon dating (Table 1).
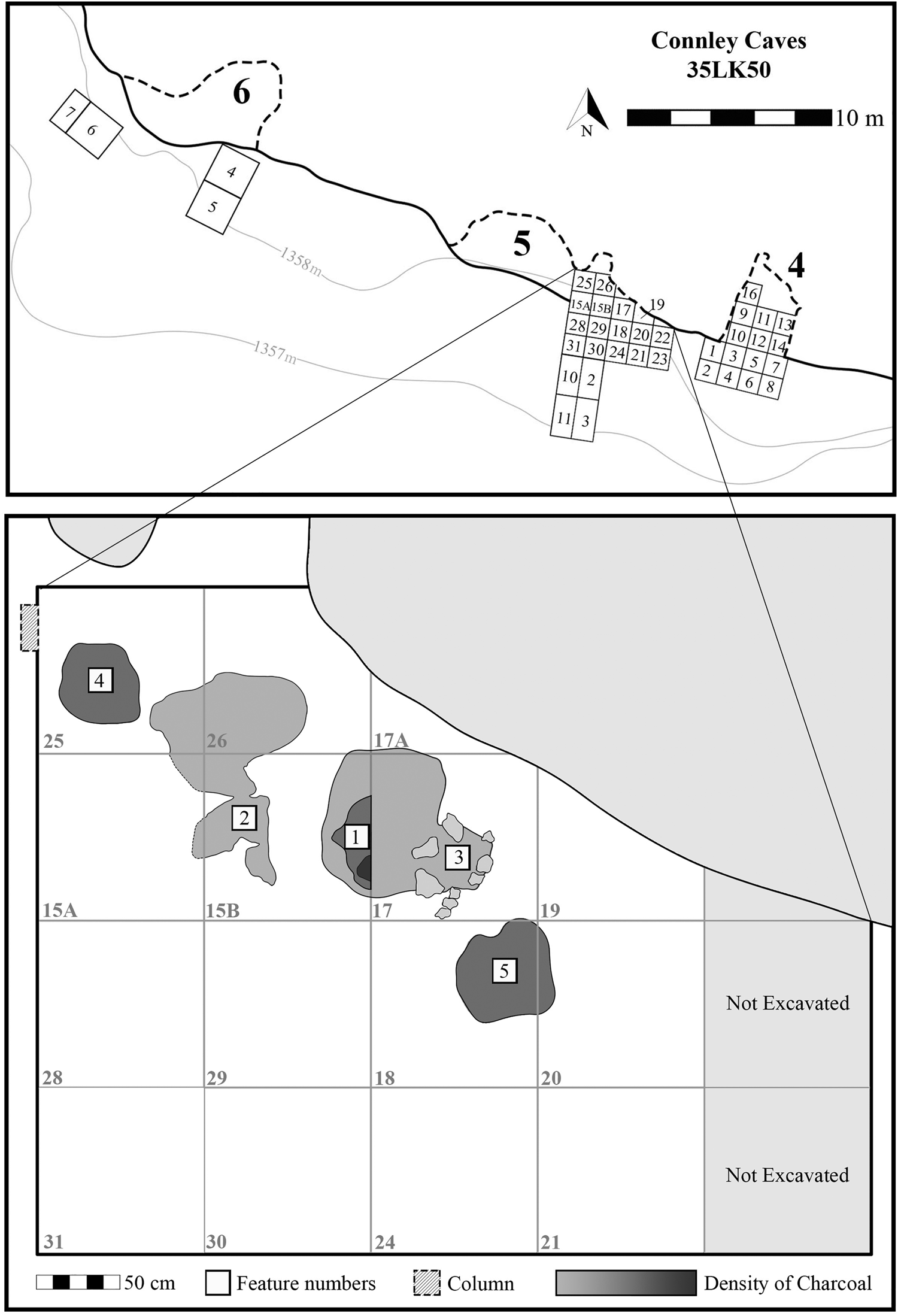
Figure 3. Planview of Connley Cave 5 (2014–2019) excavation and locations of features and column sample. Bottom image does not account for the vertical separation between features.
Table 1. Radiocarbon Ages on Artemisia Charcoal from Cultural Features in Connley Cave 5.
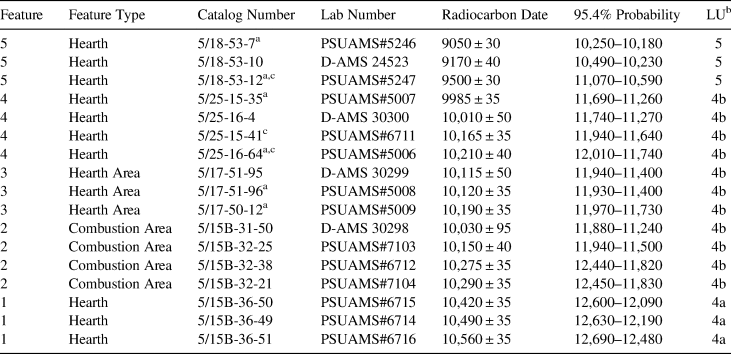
a Pretreated at the Human Paleoecology and Archaeometry Lab, University of Nevada, Reno.
b Lithostratigraphic unit.
c Considered an outlier in this study.
We processed, sorted, and analyzed the bulk sediment samples at the University of Oregon Museum of Natural and Cultural History using a dry-sieving protocol adopted from Kennedy (Reference Kennedy2018:138–139). We counted each seed fragment, including identifiable and unidentifiable fragments, as one seed in the count calculation and used soil volumes to standardize samples for comparison. To examine variation among feature and column samples, we used the exploratory statistical technique of agglomerative hierarchical cluster analysis with Ward's method and made violin plots based on seed density (average number of seeds per liter) using R 4.0.1 (R Core Team Reference Core Team2020). We computed richness, diversity, and evenness using Microsoft Excel. To evaluate whether plant remains should be attributed to human use or natural processes (e.g., animal activity or seed rain), we considered the abundance, spatial distribution, taphonomy, ecology, and ethnographic use of each taxon. We classified dietary association as “probable” when taxa had an abundance (total seed count) of five or greater and met at least three of the following criteria: (1) density (seeds per liter) is much higher within than outside features or the taxon is found only within features, (2) distance to the plant's habitat exceeds the range of rodent foraging and natural seed rain, (3) dietary use is ethnographically documented, and (4) there is evidence of burning or other nonrodent modification. Dietary association was “potential” if only two criteria were met, or if three were met but the seed count was less than five. Plants that do not fulfill two criteria are considered “unlikely” dietary constituents. The supplemental material contains further information on our materials and methods.
Results
Stratigraphy
Excavations at Connley Cave 5 revealed stratigraphy matching Connley Cave 4 (Jenkins et al. Reference Jenkins, Holcomb and McDonough2017) and clarified site-formation processes (Figures 4 and 5; Supplemental Table 2). The PHT archaeological record at Connley Cave 5 includes, from the bottom upward, windblown and colluvial deposits spanning lithostratigraphic units (LU) 2 through 5. Descriptions of each LU can be found in Supplemental Table 2. The five combustion features discussed here are preserved within LU4a, 4b, and 5. The features are distributed both horizontally and vertically throughout the Cave 5 excavation block (Tables 1 and 2). We currently interpret these features as representing separate cultural occupations, except for Features 2 and 3, which occurred at similar elevations.
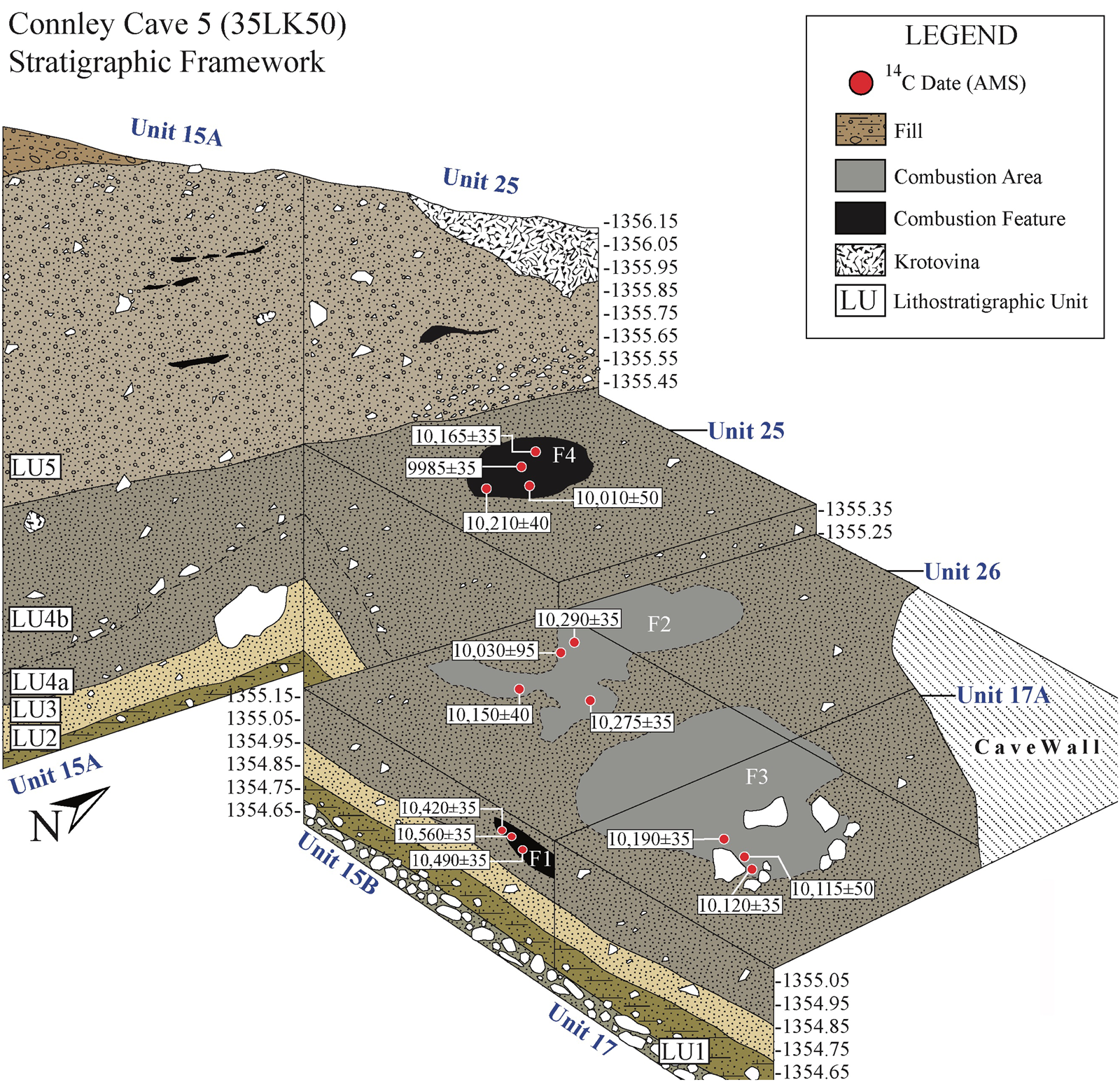
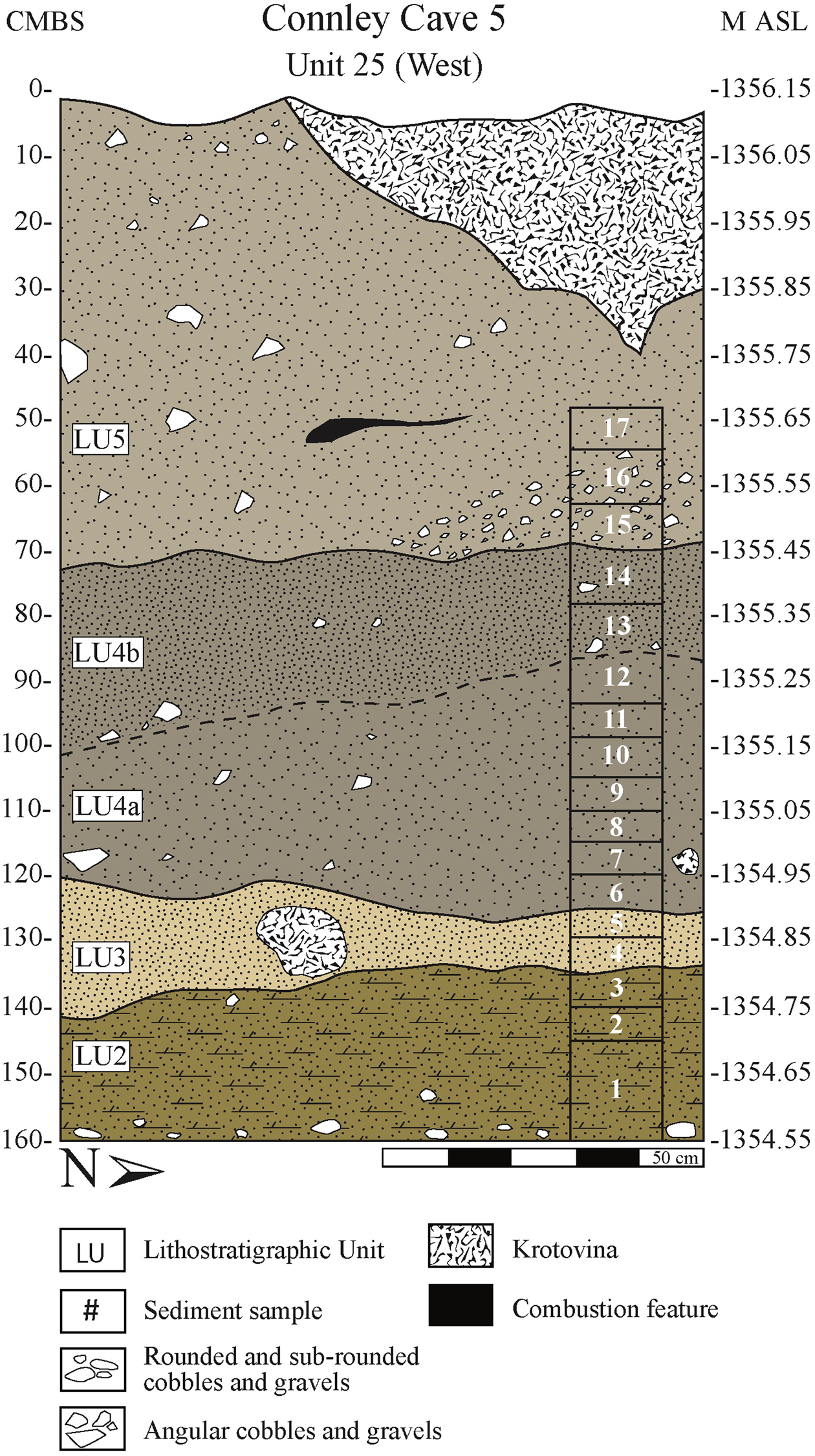
Figure 5. Stratigraphy of Unit 25's west profile with corresponding lithostratigraphic units (LUs), described in Supplemental Table 2. (Color online)
Table 2. Feature Sample Data.
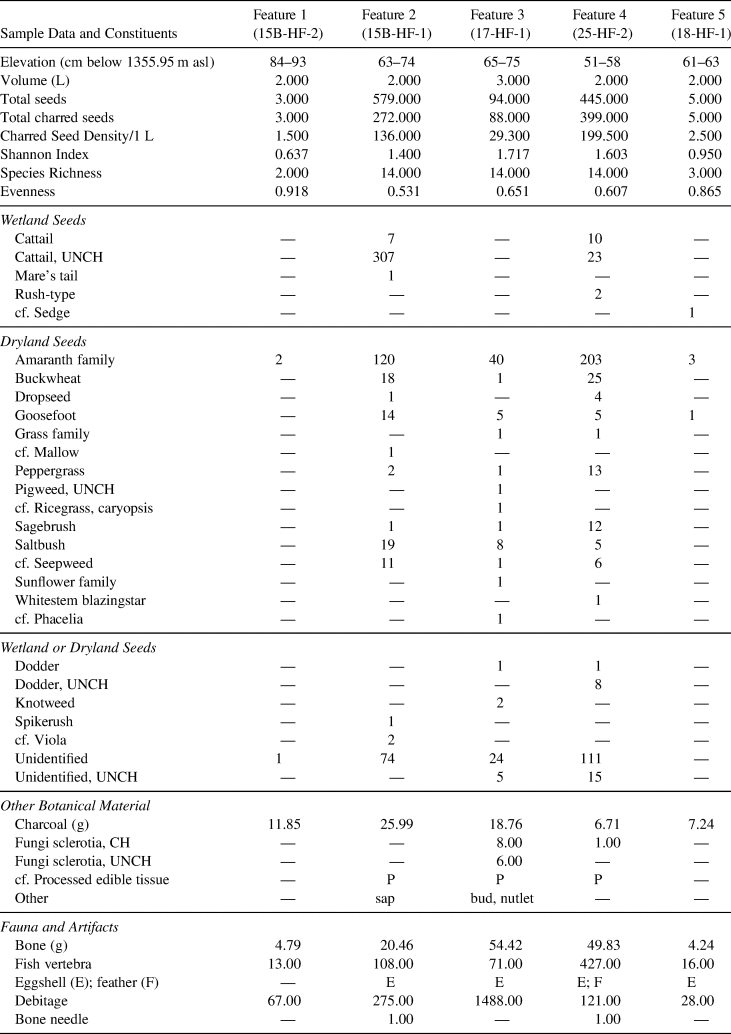
Note: All seeds are charred unless indicated as uncharred (UNCH). Values in taxa rows are total counts unless indicated as weight (g) or present (P) versus absent.
Features
Feature 1 (15B-HF-2) is a bowl-shaped hearth with intact bedding and firing structure, as indicated by stratified charcoal wood fragments and ash above rubified sediment. Feature 2 (15B-HF-1) is a combustion area characterized by irregular and widespread patches of ash, dispersed charcoal, and discolored and rubified sediment with diffuse boundaries. This combustion area may represent one or multiple hearths, a rake-out, or a combination of both that was obscured via natural (e.g., wind) or cultural (e.g., foot traffic) processes. Feature 3 (17-HF-1) is a hearth area that includes both a rock-lined hearth composed of nine cobbles arranged in a circle within Unit 17 and an area of concentrated charcoal expanding out from the rock ring toward the northwest and into Unit 15B. Similarity of the fill within the feature and the contiguous combustion area to the west suggests that some of the original contents of the hearth may have been displaced outside the rock ring via wind or human activity (i.e., a rake-out). We separately analyzed samples from within (#1781, #1994) and outside the rock ring (#2460) but consider them together as Feature 3 because their contents are likely to represent the same event. We, however, cannot rule out the possibility that these areas represent multiple events. Feature 4 (25-HF-2) is an intact bowl-shaped hearth with a rubified base located in the center of Unit 25. Feature 5 (18-HF-1) is a shallow bowl-shaped hearth in the northeast quadrant of Unit 18.
Radiocarbon Dating
Table 1 lists new radiocarbon dates on sagebrush charcoal from the five combustion features. Supplemental Text 1 and Supplemental Table 3 contain further provenience and lab information. All the dates for Features 1, 2, and 3 internally overlap at 95.4% probability. The four dates from Feature 4 fall into two groups separated by ~150 radiocarbon years. Because Feature 4 is ~20 cm higher in elevation than Feature 3, the younger two ages appear to date the feature. PSUAMS#5247 is ~400–300 years older than the other dates from Feature 5, suggesting it is the outlier. Because all dates are on charcoal from undisturbed contexts within cultural combustion features, bioturbation is not likely the cause of the variance. Dating inconsistencies are more likely explained by the burning of ancient wood from nearby woodrat middens as fuel, the reuse of old features later in time, or statistical scatter. We favor the first possibility because Features 4 and 5 are discrete hearths that are elevationally or stratigraphically younger in age than the other features. To summarize, the radiocarbon results indicate that Feature 1 dates to ~12,500 cal BP, Feature 2 to between ~12,000 and 11,800 cal BP, Feature 3 to ~11,800 cal BP, Feature 4 to ~11,500 cal BP, and Feature 5 to ~10,200 cal BP.
Archaeobotanical Analysis
Analysis of 11 sediment samples from the five combustion features yielded charred (n = 767) and uncharred seeds (n = 359; see Table 2; Figures 6 and 7; Supplemental Table 4 presents individual sample counts). Represented among the charred seeds were 21 unique plant taxa, 18 of which are known edible plants included in diets of Great Basin and California Indigenous groups (Table 3; Supplemental Table 5 presents Klamath, Modoc, and Northern Paiute plant names). Other constituents include charcoal, sclerotia (mycorrhizal fungal bodies), sap, bone, eggshell, feathers, debitage, and two bone-eyed needle fragments. Three features yielded small (<4 mm) botanical fragments displaying characteristics consistent with processed edible tissue (PET; Stenholm Reference Stenholm, Aikens and Jenkins1994) that may represent plant remains prepared as food; however, further analysis is needed to confirm this association and to identify taxonomic affiliation. Thus, we report that features contain fragments of possible PET but refrain from further interpretation. The most ubiquitous taxa (present in more than 25% of the samples) from the features are goosefoot, saltbush (Atriplex sp.), peppergrass (Lepidium sp.), seepweed (Suaeda sp.), buckwheat (Eriogonum sp.), and cattail (Typha sp.). Amaranth family and cattail seeds are in the greatest abundance (total number of seeds), followed by buckwheat and sagebrush seeds. Uncharred seeds found within features are predominantly cattail (91.9%), followed by smaller amounts of dodder (Cuscuta sp.; ~2.1%), pigweed (Amaranthus sp.; <1%), and unidentified fragments (5.6%). Charred seed density is highly variable between features, ranging from an average of 1.5 seeds per liter in Feature 1 to 199.5 seeds per liter in Feature 4. Features 2, 3, and 4 have high richness and diversity values, whereas Features 1 and 5 have low richness (Table 2).

Figure 6. Seeds (a–c, e–w) and other macrobotanicals (d, w) recovered from combustion features: (a) peppergrass; (b) cf. sedge.; (c) knotweed family (Polygonaceae) endosperm; (d) possible bud fragment; (e) phacelia; (f) mare's tail; (g) dodder; (h) spikerush.; (i) dropseed sandgrass; (j) mallow family; (k) whitestem blazingstar; (l) amaranth family; (m) goosefoot; (n) saltbush; (o-p) cf. seepweed; (q) buckwheat; (r) cattail; (s-u) sagebrush; (v) rush; (w) sclerotia. (Color online)

Figure 7. Other hearth constituents: (a–b) bone-eyed needle; (c) charred, fragmented Amaranthaceae seeds; (d) fish vertebrae (Feature 4); (e–f) examples of possible PET; (g) eggshell fragments (scale bars, 1 mm). (Color online)
Table 3. Ecological and Dietary Associations of Taxa Found in Features.
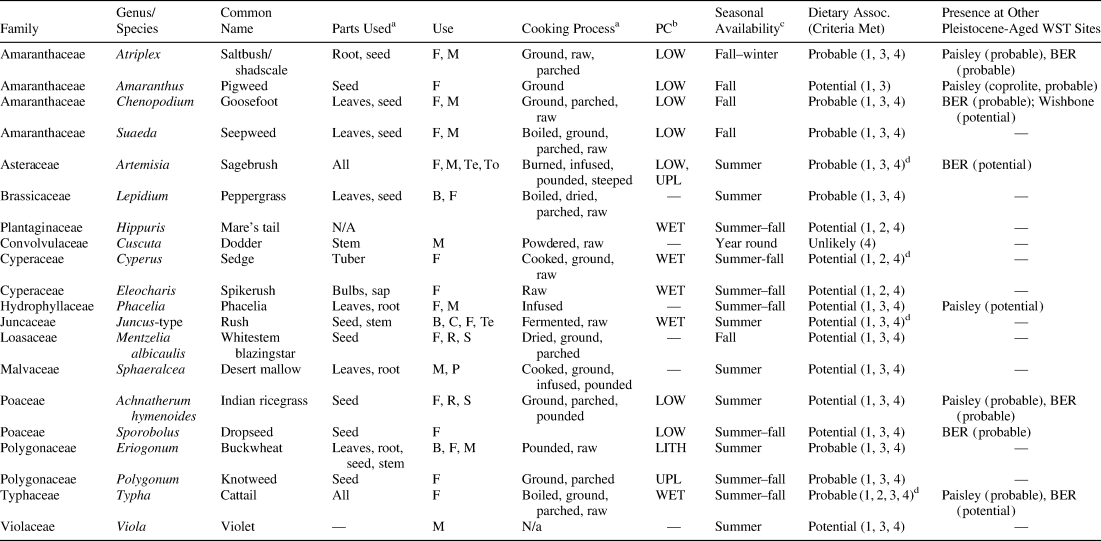
a Traditional uses and cooking process are based on ethnographic data from Indigenous groups of Oregon, Nevada, Utah, and California: beverage (B), candy (C), food (F), relish (R), sauce (S), textile (Te), and tool (To).
b Habitat classification is based on plant communities (PC) in the Fort Rock basin by Prouty (1994): wetland (WET), lowland (LOW); upland (UPL), and lithosol (LITH).
c Seasonal availability is based on data from USDA (2014) (dark grey indicates greatest availability and light grey indicates possible availability).
d Economic use could be non-dietary (e.g., fuel source or textile construction).
Column samples yielded 40 charred and 3 uncharred seeds (Supplemental Table 6). Plant taxa in the column samples include seeds of rush-type (Juncus-type), cattail, dodder, dropseed (Sporobolus sp.), goosefoot, and saltbush. Additional seeds from the grass and amaranth families could not be identified to genus. Column samples did not contain any formed tools or material resembling PET. When normalized by volume, the density of charred seeds in nonfeature samples ranged from zero to ~13 per liter (excluding sample C14, which was less than 25 cm from Feature 4, and C17, which was disturbed), suggesting that natural seed deposition is characterized by low density and low diversity.
When compared, as in the violin plots in Figure 8, there is a significant difference in the seed density of samples from Features 2, 3, and 4 (M = 109, SD = 100) and samples from Feature 1, Feature 5, and the sediment column (M = 4.6, SD = 5.8); t(4.9), p = 0.001. Cluster dendrograms identify Features 2 and 4 as outliers, whereas Features 1 and 5 group closely together. When analyzed with the column samples, Features 2 and 4 remain most dissimilar, whereas Features 1 and 5 group closely with the column samples (Figure 9). Feature 3 samples are dissimilar from the column samples, but less so than Features 2 and 4. Overall, this suggests that the archaeobotanical assemblages of Features 2, 3, and 4 are inconsistent with natural patterns of seed deposition at the site, whereas those from Features 1 and 5 are consistent with natural seed deposition.
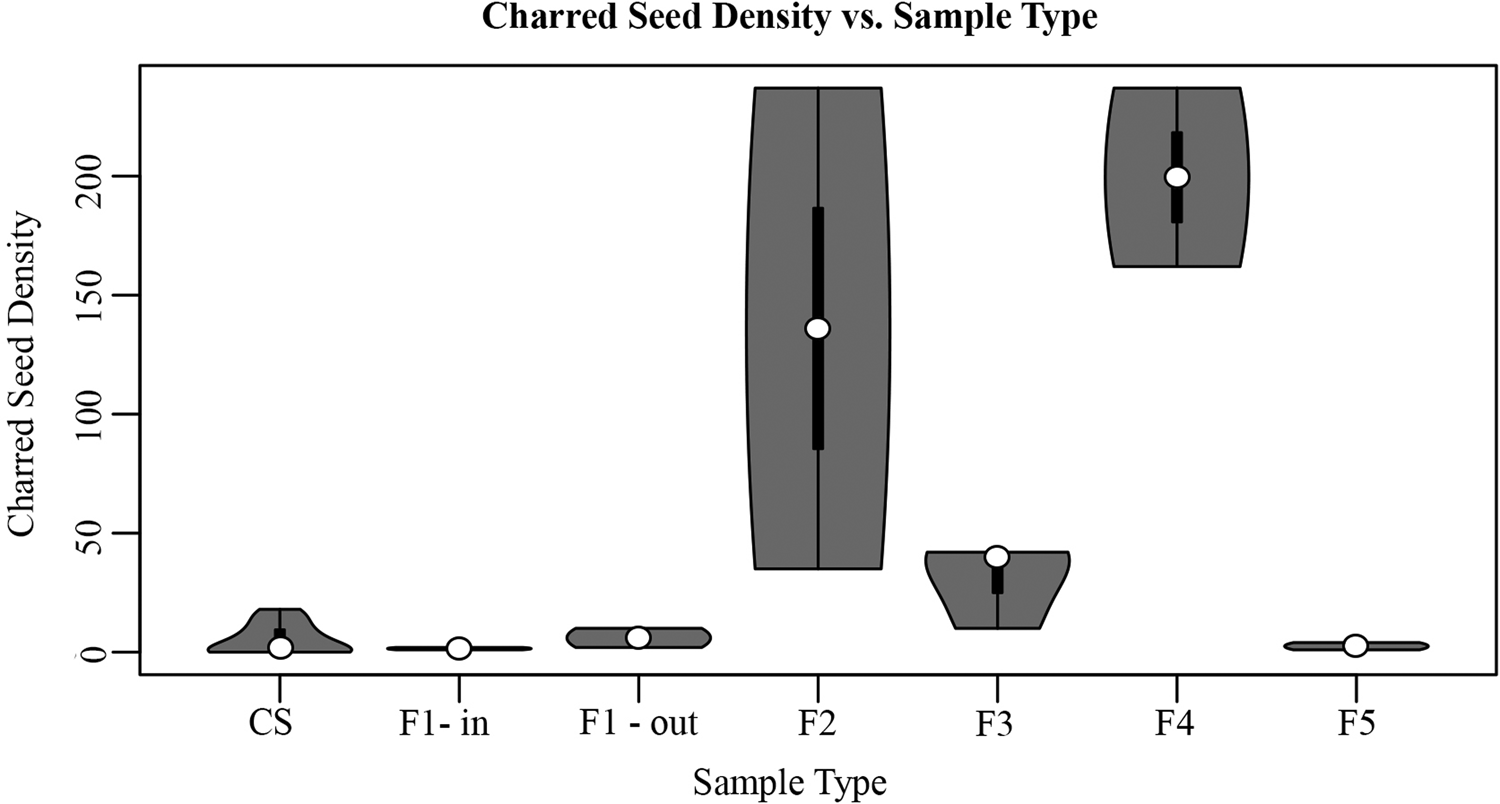
Figure 8. Total charred seed density (average number of seeds per liter) by sample type, including column samples (CS, n = 17); inside Feature 1 (F1 – in, n = 2); outside Feature 1 (FI – out, n = 2); inside Feature 2 (F2, n = 2); inside Feature 3 hearth (F3, n = 3); inside Feature 4 (F4; n = 2); and inside Feature 5 (F5, n = 2).
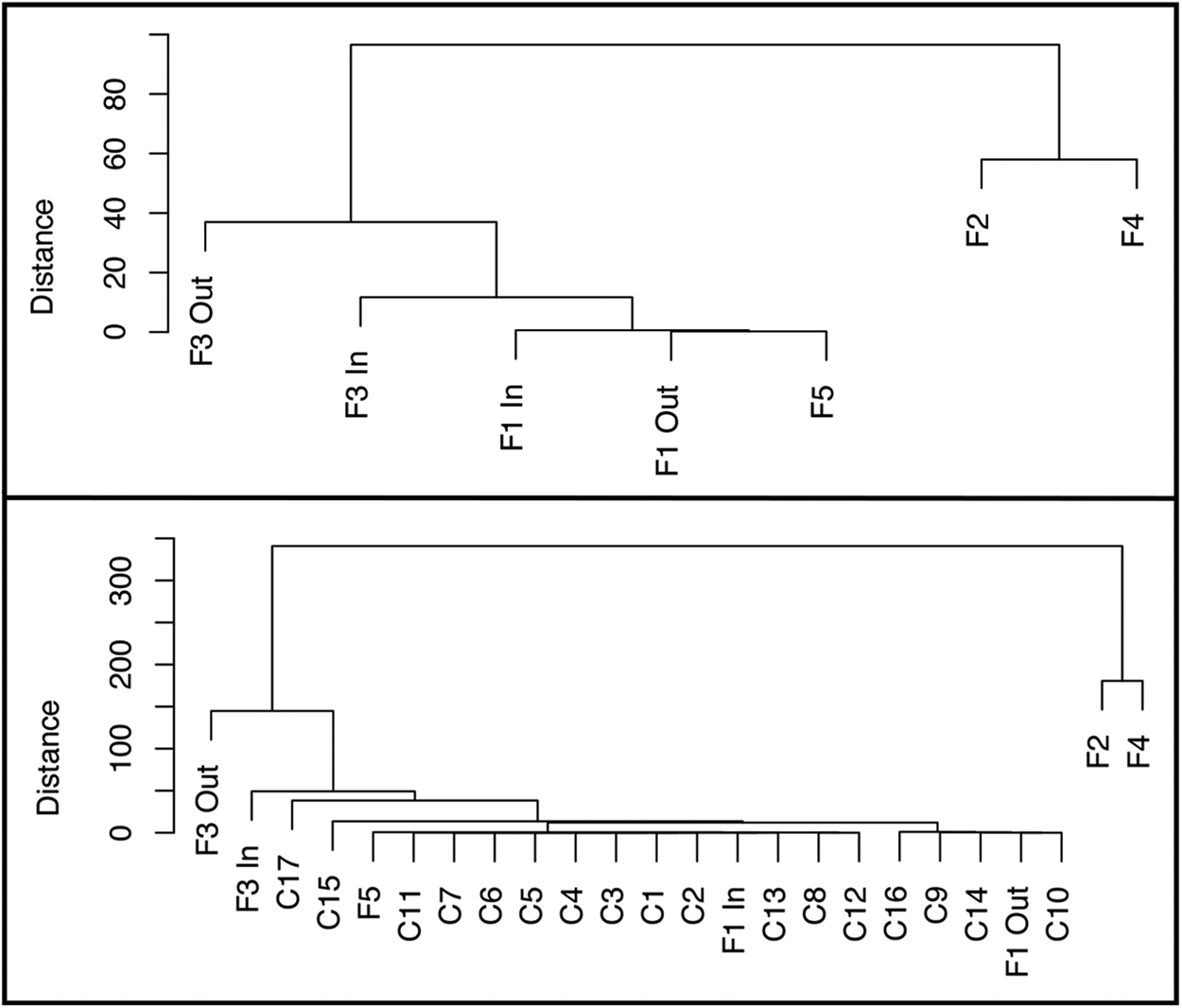
Figure 9. Cluster dendrograms of feature and column sample archaeobotanical assemblages constructed using density (average number of seeds per liter) of taxa considered “probable” and “possible” dietary constituents.
Feature Interpretations
Our analysis reveals that people visiting Connley Caves during the Younger Dryas gathered and consumed plants during multiple periods of site use. Based on our evaluative scheme, probable dietary taxa include buckwheat, cattail, goosefoot, peppergrass, saltbush, and seepweed (Table 3; Figure 10). Sagebrush seeds meet two of our criteria; however, their abundance within features is probably related to their use as fuel. Potential dietary taxa include whitestem blazingstar (Mentzelia albicaulis; partially charred), knotweed (Polygonum sp.), ricegrass (cf. Achnatherum hymenoides), phacelia (cf. Phacelia sp.), pigweed, rush, sedge, spikerush (Eleocharis sp.), violet (Viola sp.), mare's tail (Hippuris sp.), and possibly mallow (cf. Malvaceae) and sunflower (Asteraceae). Although dropseed is not commonly mentioned in the ethnographies we consulted, the seeds of this plant are consumed by Indigenous groups elsewhere in North America (Moerman Reference Moerman1998). Dropseed and several other taxa (e.g., cattail, rush, and spikerush) are also used in textile production and may represent other economic activities (Stevenson Reference Stevenson1915). In this section, we interpret each feature individually with consideration of the associated nonplant materials. A summary of artifacts associated with each feature can be found in Supplemental Text 1 and Supplemental Figure 3.

Figure 10. Summary of hearth contents (average number of seeds per liter), including all seeds recovered from features, and reflecting maximum dimensions within the feature, except for Feature 1, which was also sampled outside the hearth.
Feature 1 (~12,500 cal BP)
Feature 1 did not produce compelling evidence for dietary plant use. Densities of charred seeds inside (n = 3) and outside (n = 12) the hearth are slightly lower than the density observed in the column samples of corresponding elevation. Low density, diversity, and richness indices of plant remains in those samples suggest that they are the result of natural, rather than cultural processes. A paucity of plant remains precludes interpretation of season or foraging range; however, the lack of sagebrush seeds (the primary fuel taxa in the hearth) suggests occupation during a season other than fall. The densities of debitage, bone, and fish vertebrae are also low relative to other features.
Feature 2 (~12,000–11,800 cal BP)
The botanical remains from the Feature 2 combustion area may represent multiple activities or even occupation events, therefore providing a broader view of human behavior than assemblages from the discrete hearths. Samples from Feature 2 have high seed density, diversity, and richness measurements (Table 3). Based on their abundance and ethnographic uses, the presence of buckwheat, cattail, goosefoot, saltbush, and seepweed seeds suggests subsistence activities. The seeds of dropseed, mallow, mare's tail, sagebrush, spikerush, peppergrass, and violet may also be economic but are present in low quantities. Small fragments of possible PET are present and may represent the use of geophytes or fruits. There is a greater diversity of summer-seeding taxa but a greater abundance of fall-seeding taxa. Because the combustion area may be a palimpsest of multiple site visits, it is unclear whether people foraged in multiple habitats during a single occupation in the late summer/early fall or during multiple visits throughout the summer and fall.
Feature 2 has the highest density of charcoal of any sample and an unusually high density of fish vertebrae. The average density of fish vertebrae in Feature 2 (54 per liter) is higher than in Features 1, 3, and 5, but slightly lower than in the corresponding column sample (C13, 88 per liter), so it is currently equivocal whether they represent consumption. We also recovered the proximal fragment of a bone-eyed needle within this feature (Figure 7a). Overall, the constituents from Feature 2 show that people foraged for diverse plants in both wetland and dryland habitats and suggest that they may have also fished and sewed.
Feature 3 (~11,800 cal BP)
As a hearth area, Feature 3 provides a narrower view of behavior than Feature 2 but may also represent multiple uses or events. The rock ring demonstrates some level of hearth preparation, possibly signaling prolonged feature use. The combustion area that extends outside the rock ring could be from a rake-out or, less likely, from hearth contents displaced by aeolian processes. Analysis of the contents revealed very similar constituents within and outside the rock ring, suggesting that they reflect similar activities and possibly the same event. As such, we consider them together as one hearth area.
Feature 3 contains a high density of economic plants known from other Younger Dryas sites, such as goosefoot and saltbush seeds. Lesser amounts of buckwheat, peppergrass, phacelia, seepweed, grass, and sunflower family seeds may also reflect subsistence. Sagebrush and dodder seeds are more likely related to medicinal plant or fuel use, rather than subsistence. Both plants are important to the Paiute and Klamath for their medicinal properties (Coville Reference Coville1897; Mahar Reference Mahar1953; Murphey Reference Murphey1959; Park and Fowler Reference Park and Fowler1989; Spier Reference Spier1930; Train et al. Reference Train, Henrichs and Andrew Archer1941). Sagebrush is also a primary source of fuel at Connley Caves and is one of the many plants that dodder parasitizes (Hitchcock and Cronquist Reference Hitchcock and Cronquist2018:447). One charred nutlet fragment, two charred bud fragments, and possible PET could also be subsistence related.
All the botanical taxa in Feature 3 grow in dryland contexts, primarily in lowland habitats with minor representation of lithosol (buckwheat) plant communities. All four taxa from the amaranth family seed in fall, phacelia species seed between summer and fall, and the remaining three taxa (buckwheat, sagebrush, and peppergrass) seed in summer. Most seeds are available closer to fall, indicating a very late summer or fall occupation. Overall seed density and diversity for Feature 3 are higher than for Features 1 and 5 and most column samples, suggesting consumption, but perhaps in a more limited capacity than is reflected in Features 2 and 4. Bone, charcoal, and debitage densities are higher in the samples collected outside the rock ring than the sample within it (Supplemental Table 4). Microdebitage is high in all samples of Feature 3 and greatest (633 pieces per liter) in the northwestern quadrant of the feature area. Taken together, the contents of Feature 3 suggest that people engaged in a variety of activities, including plant gathering in dryland habitats during the late summer or early fall.
Feature 4 (~11,500 cal BP)
Feature 4 has the highest charred seed density (~200 per liter) of all samples analyzed and provides further evidence of wetland and dryland foraging. Based on abundance, we consider buckwheat, cattail, peppergrass, goosefoot, saltbush, and seepweed to be economic. The botanical assemblage is dominated by seeds of the amaranth family (49% seed total), but more than 90% of these cannot be identified beyond the family level due to high fragmentation and “popping” (see Figure 7c). This taphonomy could be the result of seed processing and parching. Additional seeds that may be subsistence related but occur in lower quantities include blazingstar, dropseed, and rush. Rush-type and blazingstar seeds are not present in any other feature samples, and the only other occurrence of dropseed is from C17. Our interpretation of dodder and sagebrush seeds in Feature 4 is the same as in Feature 3. There is a nearly even split between the number of taxa that seed in the summer versus the fall, suggesting that an occupation during late summer/early fall created Feature 4.
Feature 4 also yielded a tiny proximal fragment of a bone-eyed needle (see Figure 7b) and the lowest charcoal and highest fish vertebrae densities of any feature. The density of fish vertebrae (~213 per liter) suggests fishing activities and lends further support for wetland use. Overall, the low occurrence of debitage relative to the high density of charred seeds, fish vertebrae, and other faunal remains strongly suggests that Feature 4 is a cooking feature. The presence of both wetland and terrestrial plant taxa in the same discrete hearth indicates that cave occupants procured a range of resources from multiple habitats within a short period of time.
Feature 5 (~10,200 cal BP)
Feature 5 stands out for having low amounts of seeds and charcoal. It also has the lowest bone, fish vertebrae, and debitage densities and is the only feature without eggshell. The botanical assemblage comprises three amaranth-family seeds, one sedge seed, and one grass seed. These taxa typically seed in the very late summer or fall, but their low occurrences prohibit any strong interpretations regarding patterns of seasonality or foraging zone.
Discussion
Paleoindian Plant Use at Connley Caves
Paleoethnobotanical analysis and radiocarbon dating of cultural features from stratified contexts at Connley Cave 5 confirm that people recurrently visited the site and consumed plants throughout the Younger Dryas and very earliest Holocene. The oldest (Feature 1, ~12,500 cal BP) and youngest (Feature 5, ~10,200 cal BP) features contained few archaeobotanical remains and may represent events in which plant foods were of little importance (e.g., logistical stopovers or hunting forays) or may reflect winter or spring visits, which are harder to detect through archaeobotanical analysis. Few plants are available in the winter months, and those that are harvested in spring are less likely to be preserved (e.g., fresh shoots, leafy greens, roots). Features 2, 3, and 4 produced strong evidence for plant use but reflect different foraging habitats and variable lengths of stay. Feature 2 contained both wetland and dryland taxa indicative of variable foraging zones, although this combustion area may be a palimpsest of multiple events. As an intact hearth, Feature 4 offers a narrower view of site use and, given the co-occurrence of high-density fish vertebrae with wetland and dryland plants, suggests that visitors fished and gathered plants in multiple habitats within a short interval of time. The Feature 3 hearth area produced a range of dryland plants but lacked evidence of wetland use. Representation of both wetland and dryland resources in Features 2 and 4 may reflect divisions in labor, whereas other features may reflect converging foraging goals. Environmentally, the presence of wetland plant and animal taxa suggests that Paulina Marsh was established and productive during the Younger Dryas, whereas the absence of juniper macrofossils indicates that the vegetation surrounding the caves may have been more open than it is today.
Our first look at the archaeobotanical assemblages of these five features provides important insights on human behavior during the PHT at Connley Cave 5, but we must also recognize the limitations of this study. Due to issues of taphonomy, the botanical remains in our sampled assemblage most likely reflect a fraction of original plant use (Lee Reference Lee2012). Our view is likely biased toward plant taxa and parts that are more durable and have a greater opportunity to become discarded, carbonized, or both. Additionally, we examined just one area of a much larger site complex. Continued investigations in the site's other shelters and analysis of other dietary proxy data (e.g., zooarchaeological) will provide a more holistic picture of human activity and subsistence.
Contribution to the Paleoindian Subsistence Record
Archaeobotanical analyses of the assemblages from three Connley Cave 5 features reaffirm the inclusion of plants in the diet of Paleoindian groups and contribute new taxa to the known PHT food economy. Of the 19 taxa that we interpret as probably or potentially dietary, half have been found in other Younger Dryas-aged hearth features or coprolites in the Great Basin associated with WST technology (Table 3). These include seeds of cattail, dropseed sandgrass, goosefoot, phacelia, pigweed, ricegrass, rush, saltbush, sagebrush, and other members of the sunflower family. Seeds of the amaranth family appear in cultural contexts at Paisley Caves, Bonneville Estates Rockshelter, and Wishbone. Grass seeds have been interpreted as economic at both Bonneville Estates (dropseed and ricegrass) and Paisley (ricegrass). Hearths at those two sites contained other members of the mustard family, and a Paisley hearth contained phacelia. Rhode and Louderback (Reference Rhode, Louderback, Graf and Schmitt2007) attributed the presence of cattail seeds in hearths at Bonneville Estates to use of fluff as fire starter; however, the recovery of cattail seeds from Younger Dryas and Early Holocene coprolites at Paisley shows that dietary use should not be ruled out. Some Northern Paiute groups toast, winnow, and consume cattail seeds (Fowler Reference Fowler1992; Park and Fowler Reference Park and Fowler1989), as did people staying at Connley Caves during the Middle and Late Holocene (McDonough Reference McDonough2019). In sum, archaeobotanical data from Connley Caves and other Pleistocene-aged sites indicate that amaranths, grasses, mustards, and cattail were important and perhaps even staple plant foods for WST groups living in the Great Basin region during the late Younger Dryas and earliest Holocene.
New additions to the known Pleistocene plant-food economy include buckwheat, seepweed, and peppergrass. Buckwheat and seepweed are significant economic plants to the Klamath, Paiute, and Modoc (Chamberlin Reference Chamberlin1911; Coville Reference Coville1897; Fowler Reference Fowler1992; Kelly Reference Kelly, Kroeber, Lowie and Olson1932; Park and Fowler Reference Park and Fowler1989; Ray Reference Ray1963; Spier Reference Spier1930), and peppergrass is used by the Kawaiisu of California and other Indigenous groups throughout North America (Moerman Reference Moerman1998; Zigmond Reference Zigmond1981). Buckwheat seeds are eaten parched, ground, or raw, and other parts of the plant have medicinal value (Train et al. Reference Train, Henrichs and Andrew Archer1941). Seepweed is also a well-known plant food that has been found in other archaeological contexts throughout the Great Basin, including Middle and Late Holocene sites in the Fort Rock Basin (Cummings Reference Cummings and Minnis2004; Helzer Reference Helzer2001). Kennedy (Reference Kennedy2018) found both buckwheat and seepweed seeds in Early Holocene-aged hearths at Paisley Caves. Peppergrass is a variety of mustard that has been found in Great Basin coprolites (e.g., Hogup Cave; Fry Reference Fry1976) and hearths (e.g., Hidden Cave [Rhode Reference Rhode2003] and Camels Back Cave [Schmitt and Madsen Reference Schmitt and Madsen2005]), but this is its first appearance in late Pleistocene features.
Other possible additions to the current record of PHT plant use include blazingstar, knotweed, mallow family, spikerush, and violet. All these plants have known dietary or medicinal uses; however, except for blazingstar and knotweed, many of those uses focus on the underground or leafy parts of the plant, rather than the seeds (Chamberlin Reference Chamberlin1911; Coville Reference Coville1897; Kelly Reference Kelly, Kroeber, Lowie and Olson1932; Murphey Reference Murphey1959; Park and Fowler Reference Park and Fowler1989; Steward Reference Steward1933; Train et al. Reference Train, Henrichs and Andrew Archer1941). Some of these plants, such as mallow and violet, produce relatively few seeds compared to weedy taxa (e.g., many grass and amaranth species). Because of preparation strategies and the scale of seed production, some of these seed taxa would have had less opportunity to be introduced and carbonized within hearths. Nevertheless, all these plants have been recovered from Holocene-aged archaeological contexts in the northern Great Basin (Connolly et al. Reference Connolly, Ruiz, Jenkins and Deur2015; Dexter Reference Dexter2010; Kennedy Reference Kennedy2018; Kennedy and Smith Reference Kennedy and Smith2016; McDonough Reference McDonough2019; Prouty Reference Prouty, Jenkins, Connolly and Aikens2004; Puseman and Yost Reference Puseman and Yost2011; Sanford Reference Sanford1983; Stenholm Reference Stenholm, Aikens and Jenkins1994), and low amounts of mallow (n = 2) and rush (n = 1) seeds were found in undated cooking features in Paisley Cave 5. At least one of those Cave 5 features may have been an earth oven, based on the presence of charred starchy tissue and charred seeds of the lily (Liliaceae) family (Kennedy Reference Kennedy2018). Though they have not been directly dated, Paisley Cave 5 hearths are likely Pleistocene in age based on their stratigraphic positions. The presence of blazingstar, knotweed, mallow family, spikerush, and violet in the Connley Caves features may be related to economic use, but we consider this association as “potential” due to their low representation.
In addition to the paleoethnobotanical results, our study offers insights on other aspects of Paleoindian subsistence. Feature 4 provides the clearest evidence—including wetland plants and high density of fish vertebrae—for wetland exploitation. In the Great Basin, Blong and colleagues (Reference Blong, Adams, Sanchez, Jenkins, Bull and Shillito2020) recovered fish remains from a ~12,200 cal BP coprolite at Paisley Caves 2, and on the Snake River Plain of Idaho the Buhl Woman's isotopic signatures suggest that she consumed anadromous fishes (Green et al. Reference Green, Cochran, Fenton, Woods, Titmus, Tieszen, Davis and Miller1998; Jazwa et al. Reference Jazwa, Smith, Rosencrance, Duke and Stueber2021). Eiselt's (Reference Eiselt1997) analysis of boli from the abdominal cavity of Burial #2 at Spirit Cave, Nevada, identified abundant fish remains (n = 697) dating to 10,500 cal BP. Lastly, if the unilaterally barbed bone rod at the Lind Coulee site in Washington and the dated bone rod from Pyramid Lake (Nevada) (Dansie and Jerrems Reference Dansie, Jerrems, Lepper and Bonnichsen2005) are related to fishing (Irwin and Moody Reference Irwin and Moody1978), then there may be a growing corpus of evidence indicating that fish and fishing technology were important to WST groups.
The earliest evidence for plant consumption in temperate North America comes from Paisley Caves (Feature 2/6-4; dated to 11,005 ± 30 and 11,055 ± 35 14C BP) and Shawnee Minisink (Feature 12; dating between 11,020 ± 30 and 10,820 ± 50 14C BP). The earliest directly dated feature analyzed for plant remains at Bonneville Estates (F3.15; 10,760 ± 70 and 10,800 ± 60 14C BP) contained five charred seeds, whereas hearths dating to the mid-Younger Dryas (e.g., F4.15 dated to 10,645 ± 45 and F4.14 dated to 10,540 ± 40 14C BP) contain higher amounts and a more diverse set of charred plant remains (Goebel et al. Reference Goebel, Hockett, Rhode and Graf2021; Rhode and Louderback Reference Rhode, Louderback, Graf and Schmitt2007). Archaeobotanical data from Dust Cave, Wishbone, and other features from Paisley Caves and Bonneville Estates reflect continued plant use throughout the mid- to late Younger Dryas. Our research at Connley Caves offers a new location where people consumed plants between ~12,000 and 11,500 cal BP and adds additional taxa to the known Younger Dryas food economy.
The scarcity of groundstone technology and archaeobotanical remains from late Pleistocene sites has led some researchers to contend that plants, and small seeds specifically, did not become a significant part of the diet until the Early Holocene (Beck and Jones Reference Beck and Jones1997; Fowler Reference Fowler and D'Azevedo1986; Jennings Reference Jennings1957; O'Connell et al. Reference O'Connell, Jones, Simms, Madsen and O'Connell1982; Rhode et al. Reference Rhode, Madsen and Jones2006). The fact that when circumstances—for example, adequate preservation and sampling methods—allow, we find evidence of plant use prior to the appearance of formal tools associated with milling (e.g., groundstone) and parching (e.g., trays, basketry) raises the question of how people processed and consumed those resources. Plant taxa recovered from the Connley Caves can be prepared in a variety of ways, including being parched, ground, boiled, roasted, pounded, or eaten raw (Table 3). These preparation methods do not necessarily require specialized milling equipment. For instance, the seeds of the amaranth family can be “popped” via parching; the taphonomy of such seeds in Feature 4 (e.g., charred and split-open seed coats) suggests that people did just that. Seeds may also have been boiled in baskets or skin bags or pounded using informal stones (Coville Reference Coville1897; Kelly Reference Kelly, Kroeber, Lowie and Olson1932; Mahar Reference Mahar1953; Spier Reference Spier1930). Poor organic preservation in the lower levels of Connley Cave 5 makes the recovery of perishable artifacts unlikely, but we did recover three fragments of possible expedient groundstone in association with Features 2 and 3 that are undergoing analysis for use wear and microfossils. A recent analysis of Early and Middle Holocene artifacts at Hogup Cave in Utah found that milling stones were primarily associated with starch granules from geophytes, whereas basketry yielded starches from small-seeded plant taxa (Herzog and Lawlor Reference Herzog and Lawlor2016). Herzog and Lawlor (Reference Herzog and Lawlor2016:677) determined that their results “undercut the use of the presence of milling stones as a proxy for increased reliance on seeds” and concluded that more detailed studies are needed to understand the function of groundstone and textile tools. In sum, the lack of specialized milling equipment in early tool assemblages does not necessarily equate to an absence of seed use or the insignificance of plant foods, because Paleoindian groups may have processed plants using organic or expedient implements that are difficult to detect archaeologically.
Archaeobotanical assemblages from some of the oldest preserved hearths in North America suggest that plants have always been a part of people's diets; however, the range and intensity of plant gathering practices seem to have varied regionally and diachronically. Overall, the subsistence record associated with early WST tool technology, particularly Haskett projectile points, includes more diverse plant taxa than found in association with contemporaneous technocomplexes like Clovis, Vail-Debert, Bull Brook, and Dalton, which are dominated by a few taxa of wild nut mast and fruits (Gingerich and Kitchel Reference Gingerich, Kitchel, Smallwood and Jennings2015; Haynes and Hutson Reference Haynes, Hutson, Graf, Ketron and Waters2013; Hollenbach Reference Hollenbach, Walker and Driskell2007). The debate continues about whether these differences were driven by variability in regional resource availability, taphonomy, recovery strategies, cultural preferences, or other factors. Most likely it is a combination of causes, which require scrutiny, review, and quantification on a continental scale. To some researchers, the archaeobotanical record east of the Rocky Mountains indicates that groups opportunistically foraged low-cost, high-return plant foods—a pattern consistent with hunting-focused lifeways (Gingerich Reference Gingerich2011; Gingerich and Kitchel Reference Gingerich, Kitchel, Smallwood and Jennings2015; but see Hollenbach Reference Hollenbach, Walker and Driskell2007). Dietary data from Paisley Caves, Bonneville Estates Rockshelter, Wishbone, and now Connley Caves indicate that intentional plant gathering was part of the larger settlement-subsistence strategy for some WST groups by at least the Younger Dryas.
Implications for Settlement-Subsistence Models Based on Caloric Optimization
Anthropologists have long been interested in forager resource-acquisition decisions, and this topic has a lively history in North American Pleistocene archaeology (Grayson and Meltzer Reference Grayson and Meltzer2003, Reference Grayson and Meltzer2015; Grayson et al. Reference Grayson, Meltzer and Breslawski2021; Haynes and Hutson Reference Haynes, Hutson, Graf, Ketron and Waters2013; Surovell and Waguespack Reference Surovell, Waguespack and Haynes2009; Wolfe and Broughton Reference Wolfe and Broughton2020). Over the past half-century, human behavioral ecology (HBE) has been a productive theoretical framework for explaining past human behavior (e.g., Bettinger and Baumhoff Reference Bettinger and Baumhoff1982; Bird and O'Connell Reference Bird and O'Connell2006; Codding and Bird Reference Codding and Bird2015; Kelly Reference Kelly2013; O'Connell et al. Reference O'Connell, Jones, Simms, Madsen and O'Connell1982; Rhode Reference Rhode and Beck1999; Simms Reference Simms1987; Zeanah and Simms Reference Zeanah, Simms and Beck1999). HBE focuses on applying evolutionary theory and models of optimization to understanding human behavior within ecological contexts (Winterhalder and Smith Reference Winterhalder and Smith2000). Dietary optimization models typically use energetic returns as currency, viewing cost/benefit relationships in terms of time/calories. Though testing these models is beyond the scope of this article, botanical data from Connley Caves and other WST sites raise questions regarding the ability of caloric optimization to explain some foraging choices. We explore those issues here.
O'Connell and colleagues (Reference O'Connell, Jones, Simms, Madsen and O'Connell1982) were early advocates of the HBE approach in the Great Basin and were optimistic that with refined ecological data such models could be used to understand not only what happened in the past but also explain why it happened. Using a diet breadth model, they proposed that humans should have bypassed most low-ranked plant resources such as small seeds during the PHT due to the abundance of artiodactyl populations. Simms (Reference Simms1987) agreed that, based on energetic return rates, seeds should be ignored, but he also observed that such a prediction did not match regional ethnographic records. Pinson (Reference Pinson1999) argued that risk-adverse foragers of the late Pleistocene positioned their camps in lowland ecotonal settings where lower-ranked (i.e., small game) but predictable resources could be acquired from multiple habitats. More recently, Elston and colleagues developed a comprehensive HBE-grounded model for settlement-subsistence during the PHT (Elston and Zeanah Reference Elston and Zeanah2002; Elston et al. Reference Elston, Zeanah and Codding2014). Integrating models of optimal foraging and sexual division of labor, they proposed that groups situated their residential camps near wetlands so that women could supplement diets with small game and riparian resources while men pursued large game. They contended that the greater prevalence and productivity of wetlands during the PHT reduced the cost of travel between patches, making it more energetically efficient for groups to move to a new patch when high-ranked resources (e.g., artiodactyls) were depleted, rather than broadening their diets to include low-ranked resources. These HBE-based models have proven very useful for generating testable expectations, but they do not make specific predictions about which plant foods would be selected and when. Although there is general agreement that Great Basin groups were never strictly big-game hunting specialists, most researchers contend that foraging goals still should have focused on high-return resources, which would have been more abundant during the PHT.
Some researchers explain selection of low-ranked resources, especially seeds, based on their storability (Simms Reference Simms1985; Testart et al. Reference Testart, Forbis, Hayden, Ingold, Perlman, Pokotylo, Rowley-Conwy and Stuart1982), and this risk-reduction strategy is evident in the Fort Rock Basin by the Middle Holocene (Jenkins Reference Jenkins, Jenkins, Connolly and Melvin Aikens2004; Prouty Reference Prouty, Jenkins, Connolly and Aikens2004). Clear evidence of plant-food storage, however, has yet to be discovered in the late Pleistocene record (Smith and Barker Reference Smith and Barker2017). Storability is also unable to account for the use of highly perishable foods such as leafy greens. Therefore, neither energetic maximization nor storability/risk aversion seems to explain certain foraging choices made by some Paleoindian groups. Why then, despite comprising the lowest-ranked resource class in terms of caloric return rates (Simms Reference Simms1985), do small seeds recur at Paleoindian sites in the Great Basin?
We propose that people may have consumed plants to acquire essential nutrients needed for human growth and reproduction and that approaches from nutritional ecology may help explain those choices. Nutritional ecology examines the relationship between essential nutrient intake and health, recognizing that optimal health requires a range of micronutrients (e.g., vitamins and minerals) in addition to macronutrients, such as carbohydrates, fats, and proteins (Hockett Reference Hockett and Ollich-Castanyer2012a; Hockett and Haws Reference Hockett and Haws2003; Raubenheimer and Simpson Reference Raubenheimer and Simpson2016). This means that foragers who consume a diverse, nutrient-rich diet would benefit from a better nutritional framework for growth, reproduction, and, therefore, population increase than groups with narrow diets. Nutritional ecology has been effectively applied in the interpretation of the Pleistocene zooarchaeological record of the Great Basin (Hockett Reference Hockett, Graf and Schmitt2007), as well as in other time periods and regions (Hockett Reference Hockett2012b; Hockett and Haws Reference Hockett and Haws2003, Reference Hockett and Haws2005), but not to the archaeobotanical record of the PHT in North America. Kennedy and Smith's (Reference Kennedy and Smith2016) application to the trans-Holocene (ca. ~10,000 cal BP to contact) archaeobotanical record at LSP-1 rockshelter, Oregon, revealed correlations between nutritional diversity and population growth. Cattail and species of the amaranth and grass families from LSP-1 hearths contain a suite of essential nutrients difficult to acquire through meat alone—highlighting the potential for nutritional ecology to help understand early plant use.
Our initial comparison of available nutritional data (Phillips et al. Reference Phillips, Pehrsson, Agnew, Scheett, Follett, Lukaski and Patterson2014; USDA 2019) for taxa similar to dietary constituents recovered from Connley Caves and other Paleoindian sites shows that, overall, plant foods are the best sources for carbohydrates, folate, magnesium, Vitamin A RAE, Vitamin C, potassium, calcium, and manganese (Table 4; also see discussions in Mailer and Hale Reference Mailer and Hale2013; Phillips et al. Reference Phillips, Pehrsson, Agnew, Scheett, Follett, Lukaski and Patterson2014). Each of these nutrients is vital for a properly functioning human body and its repair, growth, and reproduction. Magnesium and folate, for example, are essential during pregnancy to support embryonic development, oocyte maturation, protein synthesis, and more; deficiencies in these and other micronutrients are linked to numerous birth complications (Fall et al. Reference Fall, Yajnik, Rao, Davies, Brown and Farrant2003; Laires et al. Reference Laires, Monteiro and Bicho2004; Ramakrishnan et al. Reference Ramakrishnan, Manjrekar, Rivera, Gonzáles-Cossío and Martorell1999). Vitamin A is needed for bone and tissue development (Brown and Noelle Reference Brown and Noelle2015), vitamin C facilitates immunological and cardiovascular functions (Chambial et al. Reference Chambial, Dwivedi, Shukla, John and Sharma2013), and potassium works as an electrolyte to regulate fluid balance, nerve signals, and muscle contractions (He and MacGregor Reference He and MacGregor2008). This initial comparison is provocative, but nutritional data for most Paleoindian plant foods do not exist, and studies show that nutritional data for similar conventional foods are not appropriate substitutes for that of traditionally gathered wild plants, as the latter may be higher in certain healthful nutrients (Burns Kraft et al. Reference Burns Kraft, Dey, Rogers, Ribnicky, Gipp, Cefalu, Raskin and Lila2008; Phillips et al. Reference Phillips, Pehrsson, Agnew, Scheett, Follett, Lukaski and Patterson2014). To fully understand the nutritional benefits of these and other Great Basin plant foods and to model Paleoindian plant use, more nutritional data are needed.
Table 4. Comparison of Macro- and Micronutrients of Traditionally Foraged Plant and Animal Taxa.
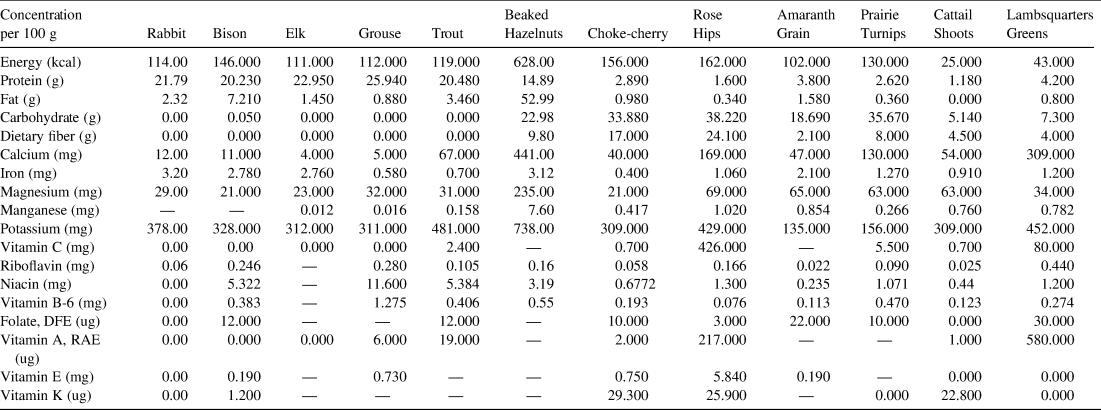
Note: Nutritional data are from the USDA National Nutrient Database for Standard Reference 28 (USDA 2019) and are given for raw food items, except for Amaranth grain, which is cooked.
We do not advocate nutritional ecology instead of HBE; rather, we suggest that integrating the two approaches may be a productive way to investigate aspects of nutrition and foraging choices. Specifically, the geometric framework for nutrition may provide a useful method for integration. The geometric framework plots variables of interest (e.g., performance consequences, lifespan, reproduction, and so on) within a geometric space defined by two or more nutritional components (e.g., carbohydrates and protein)—thereby providing a powerful graphical approach for modeling how organisms prioritize competing nutritional requirements to maximize their fitness (Raubenheimer and Simpson Reference Raubenheimer and Simpson1993; Simpson and Raubenheimer Reference Simpson and Raubenheimer1993). Applications of the geometric framework to a wide range of taxa—from slime molds to locusts, mice, and humans—have demonstrated the importance of nutrient balancing (Raubenheimer and Simpson Reference Raubenheimer and Simpson1999, Reference Raubenheimer and Simpson2016; Simpson and Raubenheimer Reference Simpson and Raubenheimer2012), leading to calls for the use of this approach to incorporate principles from nutritional and behavioral ecologies (Morehouse et al. Reference Morehouse, Raubenheimer, Kay and Bertram2020; Raubenheimer and Simpson Reference Raubenheimer and Simpson2018). It therefore follows that the application of the geometric framework to the North American Paleoindian record, and to foraging data more broadly, may help explain the increasing appearance of diverse plant resources in late Pleistocene food economies. At Connley Caves, for example, the cooccurrence of charred plant remains with a lithic toolkit including specialized projectile technology may be explained by a similar division in labor to that proposed by Elston and colleagues (Reference Elston, Zeanah and Codding2014); however, perhaps rather than directly provisioning hunting activities, plant gathering diversified the diet, providing more essential nutrients and an overall better nutritional framework for growth and reproduction.
Conclusions
This project has produced one of the most robust archaeobotanical assemblages in the late Pleistocene and earliest Holocene record of North America, revealing new aspects of foraging behavior at Connley Caves and patterns in Paleoindian subsistence more broadly. We offer these four conclusions regarding the archaeobotanical assemblages from Connley Cave 5:
(1) Analysis of five combustion features shows that people recurrently visited Connley Caves during the Younger Dryas and earliest Holocene (ca. ~12,500 to ~10,200 cal BP). Groups harvested plants during the late summer and early fall, gathering a diverse array of dryland taxa and a narrow range of wetland plants. Fishing and sewing are also indicated by some of the features’ contents.
(2) Environmental indicators, such as wetland plant taxa and fish, suggest that Paulina Marsh was established and productive in the PHT; however, marsh plants were not always the focus of food-foraging activities.
(3) The hearths from Connley Caves reaffirm and expand the diversity of plant resources in the Paleoindian food economy. These data support the proposal that WST groups in the Intermountain West had broad-based diets that incorporated resources from a variety of ecological habitats.
(4) The occurrence of seemingly low-ranked plant resources in WST sites like Connley Caves deviates from expectations of calorie-based optimization models. Our findings suggest that variables other than caloric returns may have driven some foraging choices, not only during the Holocene but also in the late Pleistocene, calling for the addition of a nutritional ecology perspective to HBE models.
Archaeobotany is essential for understanding past lifeways. Though limited, research has shown that plant remains can be recovered from a variety of late Pleistocene contexts, including rockshelter and open-air sites. Their analysis can reveal aspects of regional adaptations, seasonal mobility, foraging range, labor division, human health, and more. With concerted efforts to recover more micro- and macrobotanical remains and to integrate those data with other subsistence and environmental proxies, we will gain a better understanding of the behaviors long underrepresented in Paleoindian research.
Acknowledgments
We thank the Klamath Tribes and the Burns Paiute Tribe for the opportunity to conduct research at the Connley Caves, which are situated within their traditional territories. We also thank William Cannon and the Bureau of Land Management; the PLAYA Summer Lake Residency Program; the 2017–2019 Field School students and volunteers, especially Itzel Quiroz, Shelby Saper, and Kirsten Klement; and Brendan Culleton, Ted Goebel, Heather Thakar, Vaughn Bryant, and the National Plant Germplasm System. Funding was provided by the National Science Foundation (BCS-2054312 DDRI), Lakeview District BLM, University of Oregon Museum of Natural and Cultural History, Sven and Astrid Liljeblad Grant in Great Basin Studies, NAA/Am-Arcs Student Research Grant, Arthur Hurley, and the Texas A&M University Department of Anthropology and Center for the Study of the First Americans. We are grateful to Chantel Saban for providing the Spanish abstract translation. We thank Bryan Hockett and two anonymous reviewers for their comments, which improved this article.
Data Availability Statement
All data presented in this article are available in the supplemental material. Artifact and botanical assemblages are housed at the University of Oregon Museum of Natural and Cultural History.
Supplemental Material
For supplemental material accompanying this article, visit https://10.1017/aaq.2021.141.
Supplemental Text 1. Review of Terminal Pleistocene Archaeobotanical Data in Temperate North America and Details on Materials and Methods.
Supplemental Table 1. Feature Proveniences and Descriptions.
Supplemental Table 2. Description of Lithostratigraphic Units (LU) in Connley Cave 5.
Supplemental Table 3. Provenience and Radiocarbon Results of Feature Charcoal Samples.
Supplemental Table 4. Quantity, Density, and Diversity of Taxa Found in Connley Caves Feature Samples.
Supplemental Table 5. Names of Taxa Found in Connley Caves Features.
Supplemental Table 6. Quantity of Taxa Found in Connley Caves Column Samples.

















 Open access
Open access