



Introduction
The order Odonata in the Paleogene has become increasingly complex in recent years. It is composed not only of rather modern dragonflies (suborder Anisoptera; e.g., Archibald and Cannings Reference Archibald and Cannings2019) and damselflies (suborder Zygoptera; e.g., Nel and Paicheler Reference Nel and Paicheler1992, and see review of Florissant insects by Meyer Reference Meyer2003) but also of a growing assortment of extinct groups, such as the suborder Cephalozygoptera (found across much of the Holarctic; Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021), a species of the otherwise Mesozoic Isophlebioptera (latest Ypresian, Messel, Germany; Garrouste and Nel Reference Garrouste and Nel2015), and odonates that some authors consider belong to an undefined suborder possibly basal to the Anisoptera and Anisozygoptera (Frenguelliidae: Ypresian and Lutetian of Argentina; Petrulevičius Reference Petrulevičius2017, and references therein).
The Okanagan Highlands series of fossil localities in British Columbia, Canada and Washington, United States of America has a rich record of insect fossils from the second half of the Ypresian (early Eocene) that have been described since the late nineteenth century. Scudder (Reference Scudder1877) reported the head of a dragonfly close to the libellulid Erythrodiplax (then “Diplax”) minuscula (Rambur) from Quesnel, British Columbia, but the locality may not be in the Okanagan Highlands series. A decade later, he did not list it among British Columbia fossil insects (Scudder Reference Scudder1890), and the fossil’s odonate identity was further questioned by Handlirsch (Reference Handlirsch1910).
The first Odonata of the Okanagan Highlands were described only recently: nine dragonfly fossils of six species (five named) in five genera (four named) of Aeshnidae, one genus and species of Gomphidae, and one unnamed genus and species of an unknown dragonfly family (Archibald and Cannings Reference Archibald and Cannings2019); 16 species in four genera and one genus-level parataxon of the suborder Cephalozygoptera and the new family Whetwhetaksidae with one genus and species, which might belong to the Cephalozygoptera (Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021); and a single damselfly, a new species and genus of Euphaeidae (Archibald and Cannings Reference Archibald and Cannings2021).
The Okanagan Highlands consists of lacustrine shale, coal, and sandstone deposits scattered across about a 1000-km transect of the interior of British Columbia, Canada to just south of the international border at Republic in north–central Washington, United States of America (Archibald et al. Reference Archibald, Greenwood, Smith, Mathewes and Basinger2011). This includes the Allenby Formation of the Princeton Group in southern British Columbia, about 50 km north of the border. The formation consists of three basins in and surrounding the village of Princeton (Read Reference Read2000).
The first reported fossil insects of the Okanagan Highlands (assuming that the Quesnel locality does not belong to the series) were collected in the shale of the Allenby Formation Vermilion Bluffs Unit on “the north fork of the Similkameen River [now called the Tulameen River], and Nine-Mile Creek flowing into Whipsaw Creek, a tributary of the Similkameen…” (Scudder Reference Scudder1879, page 175B) by George Mercer Dawson in his 1877 field season (Dawson Reference Dawson and Dawson1879) and described by Samuel Scudder (Reference Scudder1879, Reference Scudder1890, Reference Scudder1895). Insects from Vermilion Bluffs shale were next collected by Lawrence Lambe along the Similkameen River in 1906 and described by Anton Handlirsch in Vienna (Reference Handlirsch1910). Since then, numerous Allenby insects were collected by W. Shaw in 1951 and by W.L. Fry in 1955, 1956, and 1957, both working for the Geological Survey of Canada. Subsequently, specimens were collected by increasing numbers of researchers, notably M.V.H. Wilson in the 1970s and W. Wehr in the 1990s. Descriptions of these insects were published (Rice Reference Rice1959, Reference Rice1967; Wilson Reference Wilson1977; Douglas and Stockey Reference Douglas and Stockey1996; Wehr Reference Wehr and Martin1998; Archibald Reference Archibald2005; Archibald and Rasnitsyn Reference Archibald and Rasnitsyn2018; Archibald et al. Reference Archibald, Rasnitsyn, Brothers and Mathewes2018), but they did not include Odonata. Here, we report the first insects of that order from the Allenby Formation.
Material and methods
We examined two fossil wings from an exposure of Allenby Formation Vermilion Bluffs Unit lacustrine shale about 4 km southwest of the village of Princeton, British Columbia, Canada. Rubino et al. (Reference Rubino, Leier, Cassel, Archibald, Foster-Baril and Barbeau2021) estimated a mid-Ypresian maximum likelihood age of 51.85 ± 0.85 Ma. old at the “Billy’s Family Restaurant” exposure of Vermilion Bluffs shale about 4 km northeast of the collecting site. We follow the systematics of Archibald et al. (Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021) and the morphological terminology used by them. Contrary character states of compared taxa are given in brackets.
Systematic Paleontology
Order Odonata Fabricius
cf . Suborder Cephalozygoptera Archibald, Cannings, and Erickson
cf . Family Dysagrionidae Cockerell
cf . Subfamily Dysagrioninae Cockerell
Allenbya Archibald and Cannings, new genus
http://zoobank.org/72f16f61-17c2-4481-bdbc-a689571cdfc6
Figure 1A–B
Diagnosis. The wing of Allenbya is most likely those of Okanagan Highlands Dysagrionidae (see discussion) but may be separated from them and from all other Dysagrionidae by space between RP1 and RP2 relatively narrow, crossvenation between them not forming recognisable IR1 [all other dysagrionid genera: IR1 distinct or at least recognisable throughout even if somewhat zig-zagged]. Furthermore, by a combination of long, strong pterostigmal brace vein present [brace vein present in some dysagrionid genera, but not as prominent]; C-RA space distal to pterostigma and RA–RP1 space three cells wide in places [more than two cells wide only in Okanagrion Archibald and Cannings, tentative dysagrionid Valerea Garrouste et al.]; pterostigma large, length about half maximum wing width (presumed width, wing damaged). Distinct from Electrophenacolestes Nel and Arillo (wings known only by basal portions) by postnodal, postsubnodal crossveins not aligned [Electrophenacolestes: more or less aligned]; by configuration of MA, MP (see below) [Electrophenacolestes: MA, MP linear, MP not far basal to MA].
Type species. Allenbya holmesae, by monotypy.
Description. As for its only species.
Etymology. The genus name is derived from “Allenby Formation”; gender, feminine.
Range and age. As for its only species.
Discussion. We believe that Allenbya is most likely a dysagrionid closely related to Okanagan Highlands genera of the family for reasons discussed below. However, as its only fossil lacks regions where character states diagnostic of that family could be assessed (Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021), a definite assignment to it (and the Cephalozygoptera) is premature. Nonetheless, we may exclude families with similar wings in preserved portions by character states such as its pterostigmal shape [Whetwhetaksidae], the density of crossvenation in, for example, the C-RA and RA-RP1 spaces distal to the pterostigma [taxa discussed below], and the configuration of MA and MP (see below) [Sieblosiidae, Frenguelliidae: MA and MP close, linear at terminations on margin; Eodysagrioninae (tentatively in Thaumatoneuridae; Huang et al. Reference Huang, Azar, Cai, Maksoud, Nel and Bechly2017): both linear but not as close near margin].
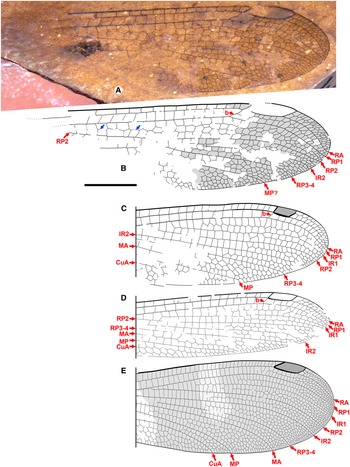
Allenbya bears a set of traits that indicate that it is closely related to the dysagrionid genera Okanagrion Archibald and Cannings, Okanopteryx Archibald and Cannings, and Stenodiafanus Archibald and Cannings (Fig. 1A–E), known only from Okanagan Highlands deposits about 150 km to the northwest and southeast, within a million years in age of the Allenby Formation, and with similar paleoclimates and forest communities.

Fig. 1. Allenbya holmesae holotype GSC 142948: A, photograph (some small darkened patches, for example, in the pterostigma, are from wetting with ethanol, not infuscation); B, drawing; C–E, distal portions of wings of Okanagan Highlands Dysagrionidae (specimens of the Stonerose Interpretive Center, Republic, Washington), modified from Archibald et al. (Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021): C, Okanopteryx jeppesenorum Archibald and Cannings holotype wing SR 13-005-013; D, Stenodiafanus westersidei Archibald and Cannings holotype SR-06-01-42; and E, Okanagrion threadgillae Archibald and Cannings holotype wing SR 98-12-10. All to scale, 5 mm. Blue arrows, see text.

Fig. 2. Aeshnidae indet. GSC 142949a & b: photographs of A, part, and B, counterpart (reversed left–right for ease of comparison); C, drawing from both as preserved, with portion (blue) folded over remainder (red); and D, wing unfolded. The darkened area posterior to the pterostigma appears to be foreign matter. All to scale, 5 mm.
The C-RA and RA-RP1 spaces three cells wide in places in Allenbya are unusual in Odonata. This occurs in the dysagrionid genus Okanagrion (three or more), the tentative dysagrionid genus Valerea Garrouste et al. (Paleocene, France: see discussion in Garrouste et al. Reference Garrouste, Wedmann, Pouillon and Nel2017 and see Archibald et al. Reference Archibald, Cannings, Erickson, Bybee and Mathewes2021), Mesozoic dragonflies of the Aeschnidiidae, and in Zygoptera in the Pseudostigmatidae, Thaumatoneura McLachlan (Thaumatoneuridae; Huang et al. Reference Huang, Azar, Cai, Maksoud, Nel and Bechly2017), Podopteryx Selys (Argiolestidae) and the Eodichromatinae, an extinct subfamily of Euphaeidae, all of which strongly differ from Allenbya in many other regards. Okanagrion (and Valerea) lacks a brace vein, present and prominent in Allenbya, and MA and CuA are linear for their entire lengths in Okanagrion (unknown in Valerea), contrary to our interpretation of these veins in Allenbya.
The blue arrows in Figure 1 indicate the basal portion of IR1 in its normal position, and the linear vein immediately posterior to it is then RP2, also positioned as expected. IR1 zig-zagged distally to become unrecognisable between a narrowly spaced RP1 and RP2 at the margin is distinct in this genus but not unreasonable.
We find it most likely that the linear, distinct vein posterior to RP3-4 is MP, not MA. If it were MA, a linear MP would have to end on the margin we believe unrealistically far basal to it. The vein MP is linear to the margin in all families compared here. As MP, it is as in Okanopteryx and Stenodiafanus (Fig. 1C–D), where MA becomes strongly zig-zagged distally and would not be recognisable in the preserved portion. Although MA would then occupy a rather narrow resulting space between RP3-4 and MP, this could be the case in Allenbya. In Okanopteryx and Stenodiafanus, CuA also becomes strongly zig-zagged distally, whereas it is linear in Okanagrion. The veins MA, MP, and CuA as in Okanopteryx and Stenodiafanus would best explain the preserved venation of Allenbya. These two genera also possess a brace vein, although it is not as prominent as in Allenbya.
Despite this evidence, we take a conservative stance and treat Allenbya as cf. Dysagrionidae and Cephalozygoptera pending a more complete specimen to clarify its placement.
Allenbya holmesae Archibald and Cannings, new species
http://zoobank.org/NomenclaturalActs/a23b896b-1274-4352-8af4-82ce2e822d60
Figure 1A–B
Diagnosis. Wing; as for genus.
Type material. GSC 142948, in the National Collection of Invertebrate and Plant Fossil Types of the Geological Survey of Canada. A partly damaged wing preserved distal to the nodus but missing much of the posterior portion from about halfway from the presumed position of the nodus to the apex. Collected by Beverley Burlingame, 19.x.2021.
Description. As in genus diagnosis, and infuscate in apical third (approximately), all preserved regions basal to that hyaline; region not preserved where oblique vein O might be if present.
Etymology. The specific epithet is a patronym formed from the surname Holmes, the maiden name of the late Ms. Dorothy Bradbeer of Agriculture Canada, mother of Beverley Burlingame, the collector and donor of the holotype, who instilled in her as a youth a love of insects and fossils.
Range and age. Vermilion Bluffs shale of the Allenby Formation, mid-Ypresian.
Suborder Anisoptera Selys
Family Aeshnidae Leach
Subfamily indet.
Genus and species indet.
Figure 2A–D
Material. GSC 142949a & b, part and counterpart, in the National Collection of Invertebrate and Plant Fossil Types of the Geological Survey of Canada. A very damaged portion of a wing, folded and missing the basal part; collected in Vermilion Bluffs shale of the Allenby Formation, mid-Ypresian, by Kathy Simpkins, v.2018.
Description. Membrane darkly infuscate at least in mid-wing, that is, basally preserved portions (perhaps absent apically by preservation); pterostigma: darkened, length about four times width; proximal, distal borders angled about 45°; anterior, posterior borders thickened; well-defined brace vein aligned with proximal border; RP2 curved with strong convex curvature just basal to pterostigma; distal portion of IR1 present, well-defined; IRP2 simple; true shape of radial planate, Rspl difficult to interpret because of damage; RP3-4 only fragmentarily preserved near terminus; MA appears gently curved, Mspl rather straight until curved posteriad near margin (but could be damaged?); medial planate appears gently widened to likely three cells wide in basal third, but could be obscured by damage; region where oblique vein O might be is not preserved. The MP–CuA space notably widens near margin.
Discussion. We assign this fossil to the Aeshnidae by its well-developed radial and medial planates and the presence of a pterostigmal brace crossvein aligned with the base of the pterostigma (Carle Reference Carle1996; Lohmann Reference Lohmann1996; von Ellenrieder Reference von Ellenrieder2002). This fossil is too incomplete and damaged to be identified below family level. However, of the Okanagan Highlands Aeshnidae, we rule out Antiquiala Archibald and Cannings and Ypshna Archibald and Cannings, by the double row of cells in the RP1–RP2 space extending more basally in the present specimen, and Eoshna Archibald and Cannings, by its shorter pterostigma. We cannot, however, exclude Idemlinea Archibald and Cannings, known only by the basal portion of a hind wing, nor the somewhat poorly preserved and unnamed Aeshnidae genus A of Archibald and Cannings (Reference Archibald and Cannings2019). It might belong to a number of extant genera.
Acknowledgements
The authors thank Kathy Simpkins (paleontology collections manager, Princeton & District Museum and Archives) and Beverley Burlingame of Princeton, British Columbia, who found the specimens described here and donated them to the Princeton & District Museum and Archives. Both have found numerous important specimens over the years in the tradition of Dawson, Lambe, Fry, Shaw, Wilson, and others and have made them available to the authors. They also thank the PDMA Board of Directors, Board President Marjorie Holland, and Museum manager Todd Davidson for the donation of the specimens from the museum to the Geological Survey of Canada, and Michelle Coyne (Curator, Geological Survey of Canada, Organic Materials Collections, Ottawa) who received them, and James Haggart (paleontologist, Geological Survey of Canada, Vancouver) for assistance. The authors thank Marlow Pellatt, of Parks Canada, for the use of a microscope and camera in his laboratory at Parks Canada, Vancouver, British Columbia. SBA thanks Rolf Mathewes of Simon Fraser University (Burnaby, British Columbia) for facilitating research. Both authors thank two anonymous reviewers for helpful comments.


 Open access
Open access