



Significant Outcomes:
-
Three sessions of conditioning are enough to promote contextual reward memory to cocaine in male and female mice;
-
Cannabidiol failed to decrease cocaine-induced conditioned place preference before the acquisition or retrieval phases of contextual memory.
Limitations
-
We studied only the acquisition and retrieval phases of conditioned place preference (CPP). More studies are needed to characterise cannabidiol (CBD) effect on CPP extinction and reinstatement;
-
We tested cocaine and CBD in both female and male mice. However, we did not investigate drug actions at specific phases of hormonal cycles.
Introduction
Cocaine use disorder (CUD) is a chronic relapsing disorder that involves compulsive cocaine-seeking despite its long-term deleterious consequences (American Psychiatric Koob & Volkow, Reference Koob and Volkow2010; Association, 2014). Repeated drug use disrupts signalling in the mesocorticolimbic pathway, including dopaminergic neurotransmission from the ventral tegmental area (VTA) to the nucleus accumbens (NAc) (Koob & Volkow, Reference Koob and Volkow2016). Other regions of importance in this disorder are the prefrontal cortex, which is involved in decision-making and impulse control; the amygdala, which is associated with emotional processing; and the hippocampus, implicated in the contextual learning forming drug-associated memories (Belin et al., Reference Belin, Jonkman, Dickinson, Robbins and Everitt2009; Kutlu & Gould, Reference Kutlu and Gould2016; Werner et al., Reference Werner, Mitra, Auerbach, Wang, Martin, Stewart, Gobira, Iida, An, Cobb, Caccamise, Salvi, Neve, Gancarz and Dietz2020). The development of drug addiction occurs in part due to environmental or contextual influences on the brain mechanisms promoting drug-seeking behaviour (Fouyssac et al., Reference Fouyssac, Puaud, Ducret, Marti-Prats, Vanhille, Ansquer, Zhang, Belin-Rauscent, Giuliano, Houeto, Everitt and Belin2021). Therefore, certain approaches to study drug addiction focus on how to prevent the acquisition or expression of contextual drug-related memories (Carey, Reference Carey2020; Asth et al., Reference Asth, Santos and Moreira2022). In studies using experimental animals, the influence of contextual stimuli associated with drug addiction can be studied with place conditioning protocols (Kuhn et al., Reference Kuhn, Kalivas and Bobadilla2019), such as the conditioned place preference (CPP) test (Bardo and Bevins, Reference Bardo and Bevins2000; US Department of Health and Human Services (HHS) 2016; Peters et al., Reference Peters, Oleson and Cheer2021).
One major neurochemical system modulating drug addiction is the endocannabinoid system, which comprises the cannabinoid receptors CB1 and CB2, endocannabinoids, and their metabolising enzymes. Cannabinoid receptors are pre-synaptically expressed in neurons projecting onto dopaminergic pathways, in the dopaminergic neurons’ cell bodies, or adjacent glial cells (Asth et al., Reference Asth, Santos and Moreira2022). The endocannabinoid system has been implicated in the initiation of substance use, in the development of compulsion and loss of behavioural control that occur during addiction (Goodman and Packard, Reference Goodman and Packard2016; De Giacomo et al., Reference De Giacomo, Ruehle, Lutz, Häring and Remmers2020; Zhang et al., Reference Zhang, Yanagida, Kamii, Wada, Domoto, Sasase, Deyama, Takarada, Hinoi, Sakimura, Yamanaka, Maejima, Mieda, Sakurai, Nishitani, Nagayasu, Kaneko, Minami and Kaneda2020; Peters et al., Reference Peters, Oleson and Cheer2021; Asth et al., Reference Asth, Santos and Moreira2022). Therefore, cannabinoids have the potential to modulate aversive and rewarding drug-associated memoy (Lee et al., Reference Lee, Bertoglio, Guimarães and Stevenson2017; Stern et al., Reference Stern, de Carvalho, Bertoglio and Takahashi2018). Cannabidiol (CBD), a phytocannabinoid, has been extensively investigated for the treatment of various psychiatric disorders, although its precise mechanism of action has remained unclear (Campos et al., Reference Campos, Moreira, Gomes, del Bel and Guimarães2012). CBD may act through multiple mechanisms, possibly enhancing endocannabinoid signalling (Elmes et al., Reference Elmes, Kaczocha, Berger, Leung, Ralph, Wang, Sweeney, Miyauchi, Tsirka, Ojima and Deutsch2015; Ibeas Bih et al., Reference Ibeas Bih, Chen, Nunn, Bazelot, Dallas and Whalley2015; Stern et al., Reference Stern, de Carvalho, Bertoglio and Takahashi2018; Galaj and Xi, Reference Galaj and Xi2019) and interacts with the serotonin 1A receptor (5-HT1A) receptor as well as with the transient receptor potential vanilloid-1 channel (TRPV1) (Resstel et al., Reference Resstel, Tavares, Lisboa, Joca, Corrêa and Guimarães2009; Galaj et al., Reference Galaj, Bi, Yang and Xi2020).
As for CBD effects on cocaine addiction, we previously reported that it prevents seizures and hepatic damage in an acute model of cocaine intoxication (Gobira et al., Reference Gobira, Vilela, Gonçalves, Santos, de Oliveira, Vieira, Aguiar, Crippa and Moreira2015). Other groups have also reported that CBD prevents certain behavioural and molecular changes promoted by cocaine in experimental animals (Luján et al., Reference Luján, Castro-Zavala, Alegre-Zurano and Valverde2018; Chesworth and Karl, Reference Chesworth and Karl2020; Gasparyan et al., Reference Gasparyan, Navarrete, Rodríguez-Arias, Miñarro and Manzanares2021; Calpe-López et al., Reference Calpe-López, Gasparyan, Navarrete, Manzanares, Miñarro and Aguilar2021; Ledesma et al., Reference Ledesma, Manzanedo and Aguilar2021) and decreases inflammatory parameters resulting from cocaine use in humans (Morissette et al., Reference Morissette, Mongeau-Pérusse, Rizkallah, Thébault, Lepage, Brissette, Bruneau, Dubreucq, Stip, Cailhier and Jutras-Aswad2021). Importantly, CBD is devoid of rewarding properties and does not induce CPP in animals (Viudez-Martínez et al., Reference Viudez-Martínez, García-Gutiérrez, Medrano-Relinque, Navarrón, Navarrete and Manzanares2019). Moreover, it has an appropriate safety profile in humans (Deiana et al., Reference Deiana, Watanabe, Yamasaki, Amada, Arthur, Fleming, Woodcock, Dorward, Pigliacampo, Close, Platt and Riedel2012; Huestis et al., Reference Huestis, Solimini, Pichini, Pacifici, Carlier and Busardò2019).
Most of these studies used male mice or rats as experimental subjects. Therefore, literature reporting cocaine and cannabinoid interventions in female animals remains scant. The few studies available suggest that both male and female mice are liable to certain cocaine effects (Hilderbrand & Lasek, Reference Hilderbrand and Lasek2014; Johnson et al., Reference Johnson, Thibeault, Lopez, Peck, Sands, Sanders, Kutlu and Calipari2019). Regarding CBD, little is known about the potential differences between males and females in terms of their responses to rewarding stimuli (Chang et al., Reference Chang, Dai and Hu2021). Recently, there has been a growing demand for pharmacological studies which include female subjects, since such approach has been largely neglected until recently, even though the development of psychiatric disorders is more common in women than in men (Altemus et al., Reference Altemus, Sarvaiya and Neill Epperson2014; Shansky, Reference Shansky2019; Radke et al., Reference Radke, Sneddon and Monroe2021). Regarding substance abuse, women may also be more vulnerable to addiction after using certain substances, especially psychostimulants (Swalve et al., Reference Swalve, Smethells and Carroll2016; Johnson et al., Reference Johnson, Thibeault, Lopez, Peck, Sands, Sanders, Kutlu and Calipari2019; Zlebnik, Reference Zlebnik2019).
Based on previous evidence on cannabinoid modulation of drug reward, we hypothesised that CBD inhibits the acquisition and retrieval of rewarding contextual memories to cocaine in the CPP paradigm. Considering the importance of including biological sex as an experimental variable, we tested CBD effects in both female and male animals exposed to cocaine. We established a 5-day CPP paradigm, with 3 cocaine-context pairing, to reduce the overall duration of the protocol.
Materials and methods
Animals
Male and female C57BL/6J wild-type mice (20–25 g), aged 8-9 weeks, were obtained from the Animal Facility of the Federal University of Minas Gerais (‘Biotério Central’). They were housed in groups of ten animals per box (41x34x16 cm) in a room under constant temperature (24 ± 2°C) and a 12 h light/dark cycle, with free access to water and food at the animal facility of the Department of Pharmacology of the Institute of Biological Sciences. The experiments were conducted during the light phase of the light/dark cycle. The authors assert that all procedures comply with the ethical standards of the relevant national and institutional guides on the care and use of laboratory animals. All procedures were approved by the Ethics Committee on the Use of Animals (CEUA) from the Federal University of Minas Gerais (UFMG), protocol number 179/2020, which is in line with Brazilian and international guidelines for the use of laboratory animals.
Drugs
CPP was induced with a dose of 15 mg/kg of cocaine (Merck & Co., Inc.) diluted in saline (NaCl 0,9%). CBD (3, 10, or 30 mg/kg) was dissolved in 5% Tween-80 in saline solution and administrated 30 minutes before the acquisition or expression phases of CPP. Cocaine and CBD was intraperitoneally administered at a volume of 10 ml/kg. The dose ranges were selected based on previous studies (Gobira et al., Reference Gobira, Vilela, Gonçalves, Santos, de Oliveira, Vieira, Aguiar, Crippa and Moreira2015, Reference Gobira, Oliveira, Gomes, da Silveira, Asth, Bastos, Batista, Issy, Okine, de Oliveira, Ribeiro, Del Bel, Aguiar, Finn and Moreira2019; Lopes et al., Reference Lopes, Bastos, Costa, Aguiar and Moreira2020).
Conditioned place preference
Cocaine-induced CPP was evaluated using an acrylic box with two compartments (15 × 12 × 12 cm each) connected by a central corridor (9.5 × 5 × 12 cm) with removable doors between each compartment (Fig. 1). The walls and floors had distinct colours and textures. One of the compartments had black walls painted with vertical white stripes and its floor consisted of equally spaced parallel stripes. The other compartment had black walls painted with horizontal white stripes and its floor consisted of a pattern of equal-sized squares. The central compartment had grey walls and an acrylic flat floor. The CPP protocol was based on previous studies (Huston et al., Reference Huston, Silva, Topic and Müller2013; Gobira et al., Reference Gobira, Oliveira, Gomes, da Silveira, Asth, Bastos, Batista, Issy, Okine, de Oliveira, Ribeiro, Del Bel, Aguiar, Finn and Moreira2019). On day 1, each animal was placed in the central compartment with free access to the box for 15 min. Animals spending more than 70% of the time in one of the compartments were excluded from the experiment (five animals showed preference for one of the compartments and thus were exclude). During the conditioning phases (days 2–4), they received injections of vehicle (morning) and, 4-5 hours later, cocaine (afternoon) immediately before exposure to a specific compartment of the box for 30 min. The allocation of each animal to one or other compartment was based on demonstrated preference on day one, with the preferred side paired with saline and the least-preferred side paired with cocaine (Huston et al., Reference Huston, Silva, Topic and Müller2013; Gobira et al., Reference Gobira, Oliveira, Gomes, da Silveira, Asth, Bastos, Batista, Issy, Okine, de Oliveira, Ribeiro, Del Bel, Aguiar, Finn and Moreira2019). Roughly, half of the animals received cocaine on each side of the CPP box (one compartment was not preferred over the other, averaged across animals). On the test day (day 5), the same procedure described for day 1 was repeated.
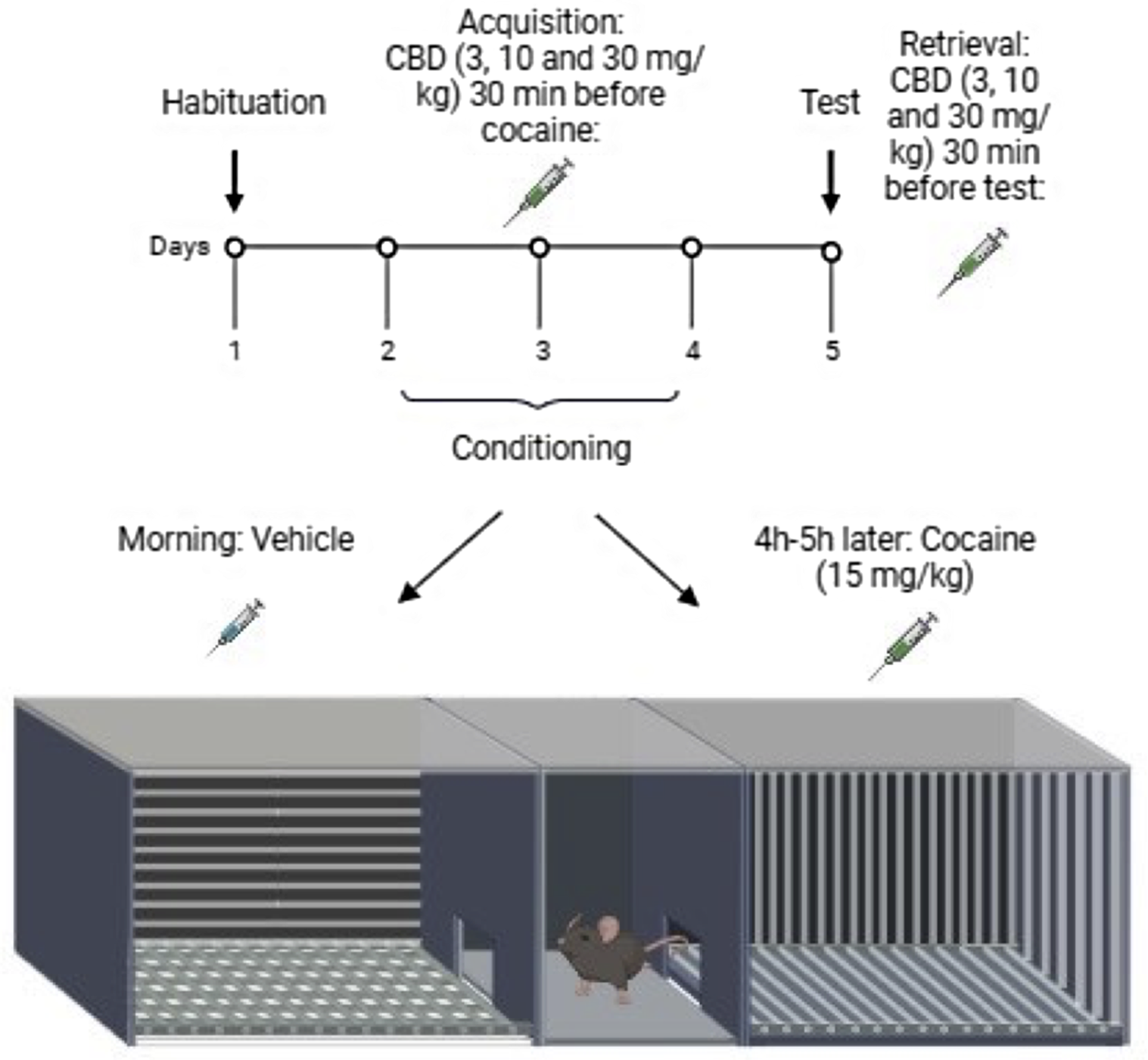
Figure 1. Protocol for testing CBD effects on the acquisition and retrieval of cocaine CPP.
The first experiment consisted of validating the 5-day CPP protocol for cocaine in male mice, with a final sample size of 8 animals receiving vehicle injection and 10 animals receiving cocaine. Next, it was tested if cocaine administration would also exert a similar effect in female animals (n = 10), as compared to males (n = 10). To test for drug effects on CPP acquisition, vehicle or CBD was administrated 30 min before each cocaine injection in the conditioning phase at the doses of 3, 10, and 30 mg/kg to both female (Veh: n = 9; CBD 3: n = 9; CBD 10: n = 10; CBD 30: n = 10) and male (Veh: n = 10; CBD 3: n = 9; CBD 10: n = 10; CBD 30: n = 9) mice. To test for drug effects on CPP retrieval, CBD was administrated 30 min before exposure on the test day to both female (Veh: n = 12; CBD 3: n = 10; CBD 10: n = 8; CBD 30: n = 11) and male (Veh: n = 10; CBD 3: n = 10; CBD 10: n = 9; CBD 30: n = 9) mice. The experiments were recorded with a video camera (Microsoft LifeCam®), and the time spent in each compartment was analysed using ANYmaze software (8th version). The data are expressed as CPP (in percentage), which is defined as the time spent in the drug-paired compartment minus the time spent in the vehicle-paired compartment *100, divided by the sum of the two sides, both for pre-test and test.
Statistical analysis
All statistical analyses and graphic elaborations were performed with GraphPad Prism 9 software. CPP scores before (‘pre-test’) and after (‘test’) cocaine conditioning were compared by repeated measures analysis of variance (RM ANOVA) followed by the Bonferroni test, when appropriate, considering the experimental groups as between factor and the pre-test and test session as within subjects. The significance level was considered as 5% (p < 0.05). Data are presented as the mean and standard error of the mean (s.e.m.) as well as individual values.
Results
Validation of the CPP protocol with male mice
Protocols for CPP differ largely across studies for some variables, including the number of drug pairings. Therefore, the validity of the CPP induction protocol for five days in male mice was verified (Fig. 2). The control group received administrations of saline during conditioning and the treatment group received cocaine (15 mg/kg) immediately before exposure to the box for three days. It was observed that cocaine induced a place preference in comparison to saline, as revealed by RM ANOVA, which detected an effect of treatment [F(1, 16) = 4.981; p = 0.0403], an effect of session [F(1,16) = 5.476; p = 0.032], and a session × treatment interaction [F(1, 16) = 8.897; p = 0.0088].

Figure 2. Cocaine (15 mg/kg) induced CPP in male mice (*p < 0.05). Data are expressed as mean ± sem (left panel), as well as individual values (right panel), analysed by RM ANOVA followed by Bonferroni test (n = 8; 10).
Validation of the CPP protocol comparing male and female mice
The validity of the protocol was verified for five days in female and male mice (Fig. 3). Both groups received cocaine, 15 mg/kg, immediately before the conditioning phase for three days. There was an overall session (pre-test vs. test) effect [F(1,18) = 27; p < 0.0001], indicating that cocaine induced a CPP, as expected. However, there was neither an effect of the variable sex [F(1, 18) = 2.602; p = 0.1242] nor an interaction between factors [F(1,18) = 1; p = 0.3164], indicating that cocaine induced CPP equally in male and female mice.

Figure 3. Cocaine (15 mg/kg) induced CPP in both male and female mice (n = 10,10, *p < 0,0001). No between-sex difference was observed. Data are expressed as mean ± sem (left panel), as well as individual values (right panel), analysed by RM ANOVA.
Effects of CBD administration (3, 10, and 30 mg/kg) on the acquisition of CPP in male and female mice
CBD was tested for its effect on the acquisition phase of place preference in males and females (Fig. 4). CBD (3, 10, and 30 mg/kg) failed to prevent the acquisition of CPP at any dose in males. There was a session effect [F (1, 34) = 55; p < 0.0001]; although no drug effect [F (3, 34) = 0,6884; p = 0,5654] or drug-session interaction [F (3, 34) = 0.1; p = 0.9430] was observed. The same pattern was observed for female animals. There was a session effect [F (1, 34) = 81.37; p < 0.0001]; although no drug effect [F (3, 34) = 0.4816; p = 0.6973] or drug-session interaction [F (3, 34) = 0.5365; p = 0.6604] was observed.

Figure 4. CBD (3, 10, and 30 mg/kg) did not inhibit the acquisition of CPP in male (n = 10, 9, 10, 9) or female (n = 9, 9, 10, 10) mice. Data are expressed as mean ± sem (left panels), as well as individual values (right panels), analysed by RM ANOVA.
Effects of CBD administration (3, 10, and 30 mg/kg) on the expression of CPP in male and female mice
We also tested the effect of CBD on the expression phase in males and females (Fig. 5). CBD (3, 10, and 30 mg/kg) failed to prevent the expression of cocaine-induced CPP at any dose, in both males and females. In the experiment with male mice, there was an effect of session [F (1, 34) = 77.83; p < 0.0001]. However, no drug effect was observed [F (3, 34) = 0.2658; p = 0.8496], the same applying to the interaction between session and drug factors [F (3, 34) = 0.2007; p = 0.8952]. Likewise, in female animals, there was an effect of session [F (3, 34) = 81; p < 0.0001], no drug effect [F (3, 34) = 0.4; p = 0.6973], and no interaction between factors [F (3, 34) = 0.5; p = 0.6604].
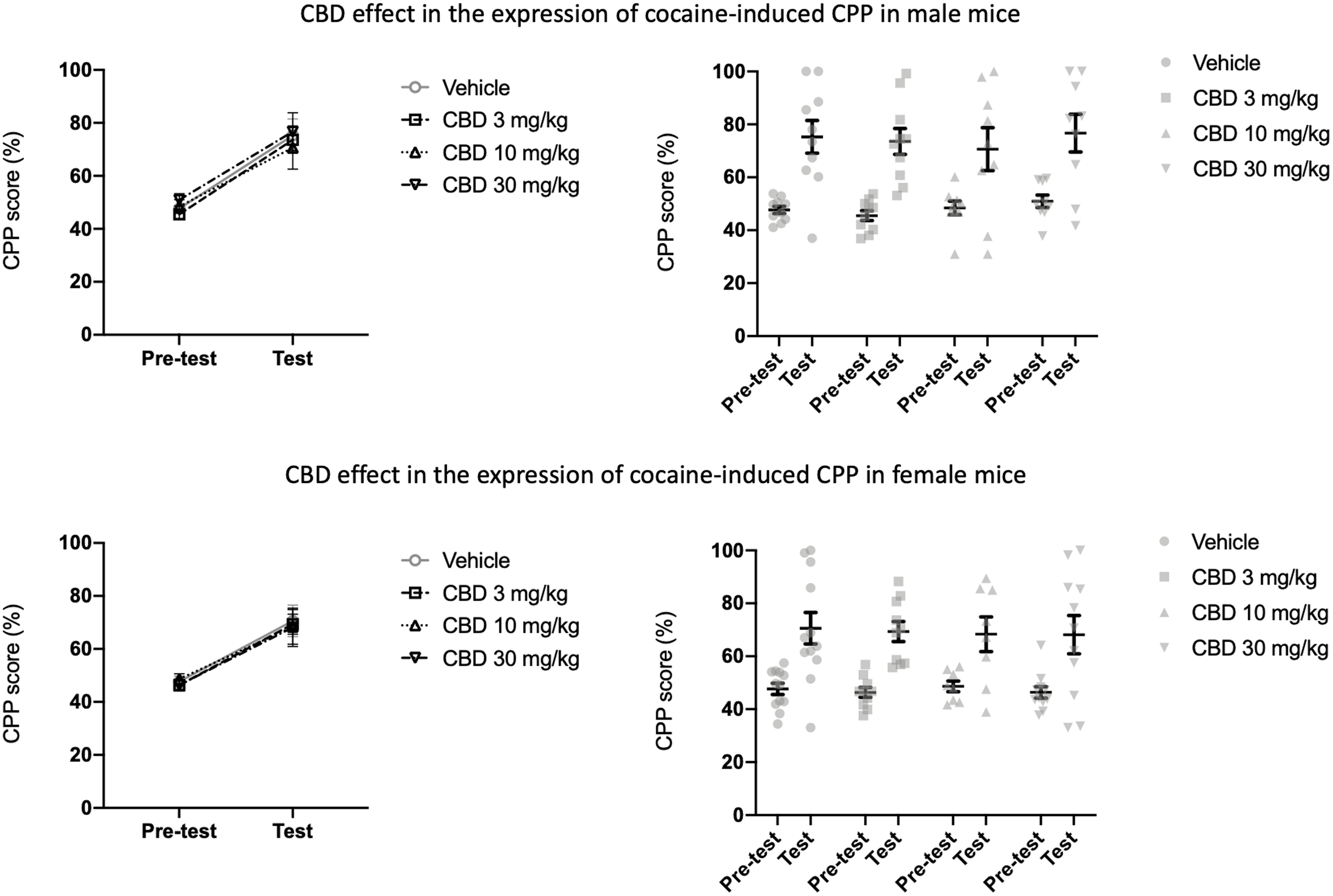
Figure 5. CBD (3, 10, and 30 mg/kg) did not inhibit the expression of CPP in male (n = 10, 10, 9, 9) or female (n = 12, 10, 8, 11) mice. Data are expressed as mean ± sem (left panels), as well as individual values (right panels), analysed by RM ANOVA.
Discussion
In this study, we showed that a 5-day CPP protocol, consisting only of three consecutive days of cocaine injections, is sufficient to induce CPP in both male and female mice of the C57BL/6J strain. This protocol complements previous studies showing the effects of cocaine with longer injection protocols (Johnson et al., Reference Johnson, Revis, Burdick and Rhodes2010; Muldoon et al., Reference Muldoon, Akinola, Schlosburg, Lichtman, Sim-Selley, Mahadevan, Cravatt and Damaj2020; Calpe-López et al., Reference Calpe-López, Gasparyan, Navarrete, Manzanares, Miñarro and Aguilar2021). We also observed that CBD failed to prevent place preference in both female and male mice when administrated on acquisition or retrieval phases.
Our initial hypothesis was based on previous evidence that cannabinoids modulate dopaminergic reward pathways (Asth et al., Reference Asth, Santos and Moreira2022). Moreover, previous studies did suggest a role for CBD in inhibiting the effects of cocaine. For instance, CBD was effective in attenuating extinction and reinstatement of cocaine-associated memories in male Wistar rats (Parker et al., Reference Parker, Burton, Sorge, Yakiwchuk and Mechoulam2004; de Carvalho and Takahashi, Reference de Carvalho and Takahashi2017). CBD also disrupted the reconsolidation of contextual cocaine- and morphine-induced contextual memories in rats (de Carvalho and Takahashi, Reference de Carvalho and Takahashi2017). In addition, CBD prevented the reinstatement of CPP induced by cocaine in male C57BL/6J mice (Ledesma et al., Reference Ledesma, Manzanedo and Aguilar2021) and male CD-1 mice (Calpe-López et al., Reference Calpe-López, Gasparyan, Navarrete, Manzanares, Miñarro and Aguilar2021). In other models to study cocaine addiction, CBD inhibited cocaine self-administration in a dose-dependent manner and attenuated brain stimulation reward in rats (Galaj et al., Reference Galaj, Bi, Yang and Xi2020).
Considering the aforementioned results, it remains unclear why CBD failed to interfere with cocaine responses in the CPP paradigm. This lack of effect could be attributed to the doses tested or the injection protocol. However, various other responses to CBD were observed at the same doses and protocols by different research groups, including ours, such as antiseizure, anxiolytic, and antidepressant-like effects (Moreira et al., Reference Moreira, Aguiar and Guimarães2006; Zanelati et al., Reference Zanelati, Biojone, Moreira, Guimarães and Joca2010; Gobira et al., Reference Gobira, Vilela, Gonçalves, Santos, de Oliveira, Vieira, Aguiar, Crippa and Moreira2015; Vilela et al., Reference Vilela, Lima, ÉB, de Miranda, Vieira É.L.M., de Oliveira, Moraes, Teixeira and Moreira2017). Moreover, other studies found that CBD failed to interfere with the acquisition and retrieval phases of contextual conditioning (Ledesma et al., Reference Ledesma, Manzanedo and Aguilar2021). Possibly, long-term administration protocols would be required for CBD to prevent cocaine-induced CPP. Indeed, CBD (10 and 20 mg/kg) attenuated cocaine effects only when administered as a pre-treatment for 10 days (Luján et al., Reference Luján, Castro-Zavala, Alegre-Zurano and Valverde2018). In another protocol, CBD reduced cocaine responses in male C57BL/6J mice, but the animals were tested 20 days after conditioning (Chesworth & Karl, Reference Chesworth and Karl2020). Finally, there is evidence that the protective effect of CBD depends on adult hippocampal neurogenesis (Luján et al., Reference Luján, Cantacorps and Valverde2019), suggesting that chronic biological processes may take place for CBD to work.
Finally, our ‘negative’ results with experimental animals have a few correlates from the clinical literature (de Meneses-Gaya et al., Reference de Meneses-Gaya, Crippa, Hallak, Miguel, Laranjeira, Bressan, Zuardi and Lacerda2021; Morissette et al., Reference Morissette, Mongeau-Pérusse, Rizkallah, Thébault, Lepage, Brissette, Bruneau, Dubreucq, Stip, Cailhier and Jutras-Aswad2021; Mongeau-Pérusse et al., Reference Mongeau-Pérusse, Brissette, Bruneau, Conrod, Dubreucq, Gazil, Stip and Jutras-Aswad2021; Rizkallah et al., Reference Rizkallah, Mongeau-Pérusse, Lamanuzzi, Castenada-Ouellet, Stip, Juteau, Brissette, Bruneau, Dubreucq and Jutras-Aswad2022). Clinical trials to investigate the effects of CBD on cocaine or crack cocaine use have yielded mainly null results for craving, withdrawal symptoms, and reinstatement. Nonetheless, in a randomised clinical trial carried out on cocaine addicts, in which the participants were treated with CBD (800 mg/kg), there was a reduction in the levels of inflammatory markers, including interleukin-6, vascular endothelial growth factor, CD14+CD16+ monocyte intermediates, and natural killer cells compared to participants receiving a placebo (Morissette et al., Reference Morissette, Mongeau-Pérusse, Rizkallah, Thébault, Lepage, Brissette, Bruneau, Dubreucq, Stip, Cailhier and Jutras-Aswad2021). Therefore, further investigation, in both human subjects and experimental animals, should clarify which cocaine-induced alterations are responsive to CBD administration.
Apart from investigating CBD activity, this study also shows that cocaine can be investigated in the CPP paradigm in a short, 5-day protocol consisting of 3 cocaine-contextual pairings. This is in contrast with most protocols in the literature, in which 3 daily, alternate vehicle or drug administrations are applied, with a total duration of 8 days (Tzschentke, Reference Tzschentke1998). Since CPP requires large sample sizes and is a laborious and long-term experiment, the use of a shorter protocol can bring advance to investigate the pharmacological modulation of cocaine responses. In addition, our results showed that this protocol is effective for testing cocaine CPP in both male and female C57BL/6J mice. Considering the importance of including both sexes as experimental variables in the biological sciences (Shansky, Reference Shansky2019), this result can serve as a reference for future studies.
In conclusion, cocaine induced place preference in both female and male mice of the C57BL/6J strain in a 5-day protocol. Regarding the effects of CBD, we evaluated the treatment before the conditioning and retrieval of the CPP responses. However, this compound failed to cocaine-induced CPP. CBD should be further evaluated for its effects on other responses to cocaine or upon CPP induced by other drugs of abuse.
Author contribution
FAM was mainly responsible for the study design. RCB was mainly responsible for conducting the experiments. All the authors have contributed to this study by participating in data analysis and writing the article. All authors have agreed on the final version of the manuscript.
Financial support
The authors thank Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) for the financial support. Project number: APQ-00741-21.
Competing interests
None.
Animal welfare ethical statement and ARRIVE guidelines
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional guides on the care and use of laboratory animals. All the protocols have been previously approved by the Committee for Ethics in the Use of Animals of the Federal University of Minas Gerais (CEUA-UFMG), which follows ARRIVE guidelines and recommendations. Protocol number: CEUA 179/2020.





