


Higher socioeconomic status (SES) in children is positively associated with cognitive attainment, among other outcomes (Duncan, Reference Duncan2012). Since J. S. Mill, Darwin and Galton, debate has raged over the contexts in which environments influence cognitive ability, culminating in the incorporation of gene–environment transactions in biopsychosocial theory (Bronfenbrenner & Ceci, Reference Bronfenbrenner and Ceci1994). Twin studies (Odintsova et al., Reference Odintsova, Willemsen, Dolan, Hottenga, Martin, Slagboom and Boomsa2018) revealed heritability around 50% for IQ in Western nations, but also moderation of endowment by SES (Tucker-Drob & Bates, Reference Tucker-Drob and Bates2016). Among impoverished US families, Turkheimer et al. (Reference Turkheimer, Haley, Waldron, D’Onofrio and Gottesman2003) found that 60% of variance in IQ was explained by the shared environment and the contribution of genes was close to 0. Among affluent families the result was reversed. Molecular genetics using non-transmitted alleles indicate that parent social competence has a large (1/3rd) influence, independent of offspring endowment (Bates, Maher et al., Reference Bates, Maher, Colodro-Conde, Medland, McAloney, Wright and Gillespie2019).
Despite most of the world living with education, industrialization, riches and democratic institutions dramatically lower than in Western countries, all existing studies of G × SES effects are based on ‘WEIRD’ (Western, educated, and from industrialized, rich, and democratic countries) data (Henrich et al., Reference Henrich, Heine and Norenzayan2010). The impact of extreme poverty on gene–environmental transaction is currently unknown, with little to no genetically informative data from people living below the low end of the SES continuum found in Western countries. To address this gap, we present data from public schools in West Africa where child mortality is 30 times the Western average (World Bank, 2016), just 40% of the population are connected to the energy grid, and 70% of the sample live on a family income of less than $1.25 a day, attending schools where education expenditures are just 16% of the levels recommend by UNESCO, even when prorated to GDP (Uche et al., Reference Uche, Ihugba and Nwosu2013). Students in the sample suffered multiple deprivation, from absence of electric lighting to overcrowding in classrooms, lack of school facilities, extending to limited access to toilet facilities and clean drinking water, poor sanitization, and undernutrition. Severe funding limitations mean public school teachers are often unqualified and schools are often closed. Compounding these issues, large average family sizes and limited financial resources incentivize child labor, which compete with learning activities. Previous studies have reliably associated such environmental characteristics with poor learning outcomes and cognitive development (Boivin et al., Reference Boivin, Giordani, Ndanga, Maky, Manzeki, Ngunu and Muamba1993). Their biopsychosocial effect, however, has not been assessed.
Methods
Subjects
A total of 3192 twin individuals attending 272 public secondary schools covering the poorest 5 of 6 school districts in Lagos State, Nigeria participated (314 monozygotic male [MZM], 620 dizygotic male [DZM], 452 monozygotic female [MZF], 764 dizygotic female [DZF] and 1042 opposite-sex dizygotic [OSDZ]). The mean age of the twins was 14.6 years (SD 1.7 years). Data collection occurred during 2013 and 2014. For same-sex twins, zygosity was determined using 18 DNA microsatellite markers. Our research was approved by the Health Research and Ethics Committee of the Lagos State University Teaching Hospital and carried out in accordance with the provisions of the World Medical Association Declaration of Helsinki.
Measures
Ability was assessed using the Raven (Reference Raven2008) Standard Progressive Matrices Plus (SPM) and Set B of the Mill Hill Vocabulary test (MHV; Raven, Reference Raven2008), two widely validated measures of ability. The SPM consists of 60 items of nonverbal stimuli that include visual matrix of patterns and shapes. Each matrix has one piece missing. Participants are requested to choose the missing pattern from a choice of six or eight. The 60 items (matrices) are organized into 5 sets (A, B, C, D and E) constructed to become increasingly difficult moving from A to E. The MHV consists of 44 multiple-choice questions listed in order of difficulty that ask for a synonym of a word. We encouraged twins to complete all items of the SPM and MHV, asking them to make their best guess when they felt items were very difficult. We did not limit the testing time so that twins could show their maximum performance. For SPM and MHV, one point was assigned for each correct answer; the skipped items or wrong answers received no point without deduction.
A parallel analysis suggested the ability scales were well modeled as a single factor (eigenvalue 2.92), and we formally characterized general cognitive ability as each subject’s score on the first unrotated component of the five SPM scales and the MHV.
SES was assessed using parental educational attainment, household assets (such as a car, air conditioner, TV, refrigerator or home computer) and number of children living in the home (as large family sizes are associated with lower wealth and fewer resources per child). These were administered to each co-twin individually and averaged across co-twins for reliability. We also recorded whether or not the children currently worked for money. SES was assessed as scores on the first principle component of a factor analysis of the three SES measures, and each family was scored on this factor. Family cohesion and chaos were assessed using the Family Adaptability and Cohesion Evaluation Scale (FACES II) (Olson et al., Reference Olson, Portner and Bell1982). Cohesion assesses emotional bonding between family members, whereas flexibility (chaos) is a dimension of change in family leadership, roles and rules, with high scores reflecting chaotic family relationships.
Procedures
Twins were tested in small groups either in the school library or in a special classroom. To prevent possible cheating on the test, members of a twin-pair were strictly monitored to sit apart. During the testing, twins were instructed to complete a questionnaire booklet where the contents were in the order of SES questions, SPM, MHV and other psychological questionnaires, including FACES II. A local research assistant explained how to solve the questions in the SPM and MHV with sample items. Twins were not allowed to move to the psychological questionnaires until they finished the SPM and MHV. No time limit was applied, but the whole protocol typically took approximately 2½ h. A minimum of one research assistant and the first author (YMH) were present in the testing room to give instructions and supervise testing. Typically, however, teachers and additional research assistants were available to help the testing. The distribution of age, SES, and scaled MHV and SPM scores are shown in Figure 1.
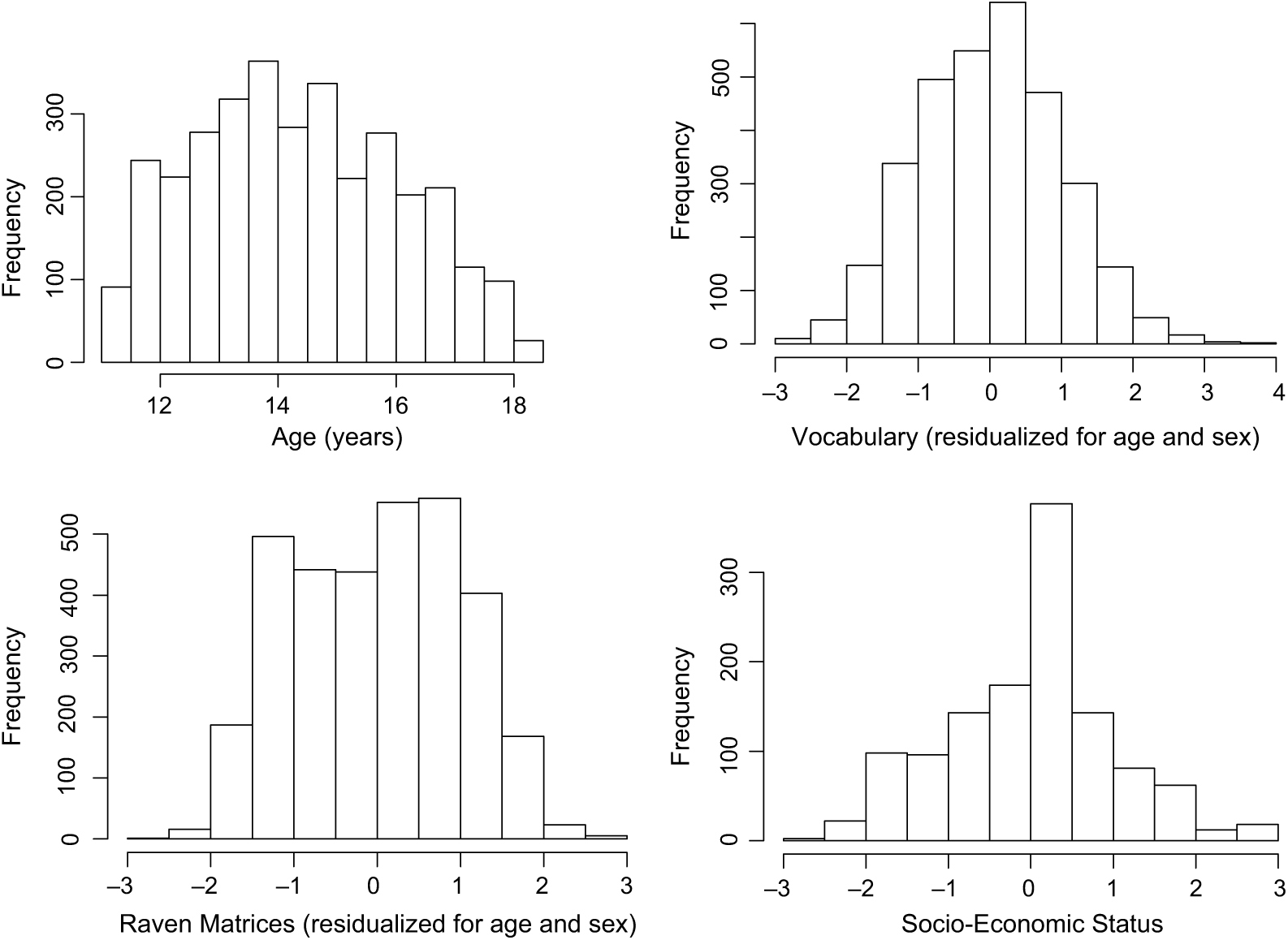
Fig. 1. Distributions of age, SES, scaled vocabulary (MHV) and Raven (SPM) scores.
Analyses
Prior to testing the gene–environment interaction model (which was our main focus), twin correlations were computed and a scalar sex-limitation model (Bates, Maes et al., Reference Bates, Maes and Neale2019) performed to determine the appropriateness of aggregating the sexes into monozygotic (MZ) and dizygotic (DZ) groups. In line with previous analyses of IQ data, sex-limitation models indicated that additive genetic, shared environment and nonshared environment variances did not significantly differ across sexes. As assortative mating for educational level can generate spurious estimates of shared environmental influences on education-related measures such as IQ, we computed a correlation for the highest level of education the twins’ mothers and fathers completed in the current sample. Our spousal correlation of .65 was very similar to those reported in studies conducted in other countries in the world (e.g., Heath et al., Reference Heath, Eaves, Nance and Corey1987; Hur, Reference Hur2003; Reynolds et al., Reference Reynolds, Baker and Pedersen2000; Torche, Reference Torche2010) and a prior study based on parents of a large (N > 7000) sample of singleton students in Lagos State in Nigeria (Hur, Reference Hur2016). This can theoretically generate artefactual G × SES (Loehlin et al., Reference Loehlin, Harden and Turkheimer2009). Power to detect G × SES was assessed using the umxGxE and umx_makeTwinData functions to generate 1000 instances of a G × SES model with our sample size and the additive genetic influence on IQ moving from .2 at −3 SDs of SES to .65 at +3 SDs of SES (matching the meta-analytic aˋ of 0.06 reported by Tucker-Drob & Bates, Reference Tucker-Drob and Bates2016), with C fixed at 0.2, and E at 0.4. To focus power on the main hypothesis — that the moderation effect of SES on observed genetic variance (aˋ) would be significant — a true model was estimated with freely estimated A, C and E as well as free aˋ and linear and quadratic mean effects of SES. This was compared with a 1-df test against a null model in which aˋ was fixed at 0. The p value for this model comparison was obtained using umxCompare and accumulated across 1000 replications. This yielded a power estimate (the proportion of replicates yielding a significant difference between the models) of 0.814. Genetic influence on IQ was estimated in a classical ACE model, decomposing variance in IQ in the twins to the influence of latent additive genetic (A) effects, correlating either 1.0 for monozygotic or .5 for dizygotic twins, shared environmental (C), correlating 1.0 across members of a twin pair regardless of zygosity, and unique environmental (E), influences specific to each twin including measurement error. G × SES was assessed in a Purcell (Reference Purcell2002) model, in which mean effects are modeled as a function of both average effects plus effects of the moderator, and variance in the trait (here IQ) modeled by the latent A, C and E variables, each taking an additional parameter, moderated by SES (see Figure 1).

Fig. 2. Genetic (a), Shared environment (c) and Unique environment (e) effects and SES moderation effects (aˋ, cˋ and eˋ, respectively) in full G x SES model. 95% CI in brackets: significant paths are shown in bold type.
Results
The measures of ability had adequate reliability (Cronbach’s alpha .88 for the SPM and .66 for MHV). These values are somewhat below those of the UK standardization sample (split-half reliability of .94 for the SPM; Raven, Reference Raven2008). However, replicating their relationship in normative samples (Raven, Reference Raven2008), the correlation between scores on the SPM and MHV was r = .40. Age was modestly but significantly correlated with SPM and MHV (r = .12 with SPM, r = .15 with MHV). While SPM was significantly higher in males than in females, the opposite was true for MHV. However, these sex differences were not large in any of the measures (d = 0.25 for SPM; d = 0.13 for MHV). Table 1 shows age- and sex-corrected correlations among SPM, MHV, SES, Family Cohesion and Chaos in our sample.
Table 1. Age- and sex-corrected correlations among ability measures, SES and family chaos (reverse scored) and cohesion
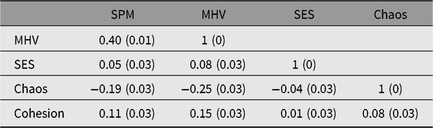
Note: SPM, Standard Progressive Matrices Plus; MHV, Mill Hill vocabulary; SES, socioeconomic status.
Standard errors are in parenthesis.
Twin correlations (Table 2) suggested heritable variance in ability in the sample (MZ correlation exceeded DZ correlation), but also evidence of shared environment effects (DZ correlation exceeded ½ the MZ correlation). Similar correlations obtained for the male and female MZ and DZ pairs, suggesting no sex limitation: male MZ and DZ pair age-corrected correlations for SPM were .57 (.05) and .37 (.05), respectively. For female MZ and DZ pairs, these correlations were .55 (.05) and .47 (.04), respectively. Opposite-sex DZ pairs correlated .38 (.04) for SPM. Male MZ and DZ pair age-corrected correlations for MHV were .70 (.04) and .44 (.05), respectively, and female MZ and DZ pair age-corrected correlations were .64 (.04) and .50 (.04), respectively. Opposite-sex DZ pairs correlated .46 (.04) for MHV.
Table 2. Age-corrected twin correlations for Standard Progressive Matrices Plus (SPM) and Mill Hill Vocabulary test (MHV)
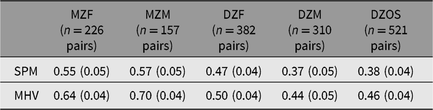
Notes: MZF, female monozygotic twins; MZM, male monozygotic twins; DZF, female dizygotic twins; DZM, male dizygotic twins; DZOS, opposite-sex dizygotic twins.
Standard errors are in parenthesis.
Heritability of Cognitive Ability Under Extreme Deprivation
As the entire sample forms what would in a Western dataset comprise an extreme deprivation group, we began by examining genetic and environmental influences in the sample as a whole, with no moderation by SES. This allowed us to test the prediction that, in extreme poverty, heritability effects would be negligible, with shared environmental factors and unique environment both having large effects accounting for the predominate fraction of differences in cognitive abilities (Toto et al., Reference Toto, Piffer, Khaleefa, Bader, Bakhiet, Lynn and Essa2018). For the SPM, however, parameter estimates were surprisingly large and significant .31 for additive genetic variance (95% CI [.16, .45]), and only slightly smaller, and significant .25 for shared environment (95% CI [.15, .36]). Unique influences were estimated at .44 (95% CI [.38, .50]). Estimates for MHV were comparable, with slightly higher heritability (A = .40, 95% CI [.27, 52]; C = .28, 95% CI [.18, 37]; E = .32, 95% CI [28, 38]). The non-moderated data, then, were not compatible with the hypothesis that in extreme poverty genetic influence on cognitive ability drops to near 0. Nor was evidence found for elevated shared environment effects. Instead, both heritability and shared environment levels at the population level lie within the range of moderate values found in Western teenage twin samples (Tucker-Drob & Briley, Reference Tucker-Drob and Briley2014).
Testing G × SES Moderation of Heritable Influence on Cognitive Ability
We next moved to test evidence of SES interaction effects (Purcell, Reference Purcell2002). To formally test for SES moderation, we used the Purcell (Reference Purcell2002) parameterization to fit G × SES interaction models to the data. This model includes effects of the moderator on the means of the dependent variable, a precondition for valid testing of SES moderation of genetic and environmental variances.
As seen in Table 3, neither aˋ nor cˋ (the genetic and shared environmental moderator terms, respectively) showed substantive effects, and could be dropped without significant loss of model fit, suggesting that SES differences were not related to expression of genetic effects on cognitive abilities in this sample. Results were similar for analyses focused on separate verbal (MHV) or spatial reasoning (SPM) ability, with all moderation able to be dropped without significant loss of model fit, χ²(3) = 3.83, p = .28 and χ²(3) = 0.287, p = .962, respectively. In follow-up analyses, we explored the role of child labor and of household chaos (Hart et al., Reference Hart, Petrill, Deckard and Thompson2007) as moderators of revealed ability. Family chaos was significantly negatively related to cognitive scores (r = −0.23, 95% CI [−0.28, −0.19], t(1599) = −9.53, p ≤ .001 in twin 1). Dropping moderation on C for family chaos lead to a significant loss of fit, χ²(1) = 4.458, p = .035. The preferred model for family chaos moderation dropped moderation on A and E, preserving moderation on C, χ²(2) = 0.31, p = .857, (compared to the full model). Family cohesion showed no evidence of moderation of ability, χ²(3) = 3.927, p = .269, in comparison with the full moderated model.
Table 3. Model fits testing genetic and environmental influences on general cognitive ability and moderation by SES
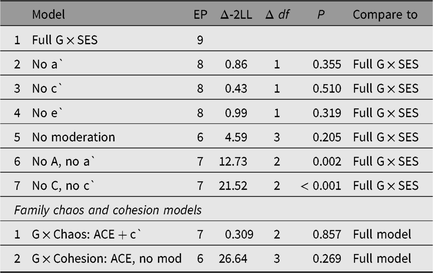
Notes: A, additive genetic effects; C, shared environmental effects; E, nonshared environment and measurement error; EP, estimated parameters; mod, moderation.
aˋ, cˋ and eˋ = moderator effects for A, C and E, respectively.
Overall, the results confirmed conventional associations of chaos and SES with offspring cognitive ability, strong support for heritable and shared environment effects on cognitive ability, a beta weight for aˋ similar to that reported for the US sample, but nonsignificant here, and family chaos moderation of shared environment effects on cognitive ability.
Discussion
Our study reports four main findings. First, both genes and shared environment played strong and significant roles in individual differences in cognitive abilities in extreme poverty. Second, levels of both genes and shared environmental influences on cognitive abilities in extreme poverty were similar to those found in much wealthier ‘WEIRD’ Western twin studies. Somewhat higher estimates of unique environment as compared to Western data probably reflected higher levels of measurement error in the current sample because we forced children to guess difficult items. As indicated above, reliabilities were somewhat lower in current sample as compared to those found in the UK standardization sample. Third, SES moderation of genetic influence on cognitive abilities was weak, but compatible with US data. The hypothesis that a critical poverty threshold exists for genes associated with cognitive ability to generate variance in attained ability was not supported. Data are compatible with canalization and resilience models of cognitive development. This is not to say, of course, that there are not mean level effects and/or effects on domains such as health. Fourth, shared environmental variance in cognitive ability was moderated by family chaos. Additional work is required to identify the nature of mechanisms coupling developmental outcomes to family environment, for instance, SES moderation of effects of interests (Zheng et al., Reference Zheng, Tucker-Drob and Briley2019). Replications are desirable, and in particular, we would be interested to study replication and mechanism of the moderation of shared environmental variance by family dysfunction.
These findings should inform thinking regarding the role of SES in cognitive development at a global level and motivate additional studies in the large swathes of global diversity excluded from studies of cognitive development to date (Toto et al., Reference Toto, Piffer, Khaleefa, Bader, Bakhiet, Lynn and Essa2018). The present data suggest that extreme inadequate environments including minimal teacher efficacy and low community literacy (Smith-Greenaway, Reference Smith-Greenaway2013) may create environments which reveal individual differences in ability to learn despite such poor opportunity.
Our study has limitations as well as strengths. It will be important for future research to study other ages and other measures of IQ and SES (Duncan, Reference Duncan2012). We hope researchers developing interventions for those in absolute poverty incorporate genetics and families in their studies. As the first study of gene × SES interaction effects to include a large sample of twins living in extreme poverty, our results suggest that causal influences on cognitive development under conditions of extreme poverty may be similar to those in average environment in many western developed countries.
Acknowledgments
This study was funded mainly by Ulster Institute for Social Research (UK). We would like to thank twins who participated in this research.






 Open access
Open access