


Numerous heritability studies show that brain structure is under moderately strong genetic control (Kremen et al., Reference Kremen, Prom-Wormley, Panizzon, Eyler, Fischl, Neale, Franz, Lyons, Pacheco, Perry, Stevens, Schmitt, Grant, Seidman, Thermenos, Tsuang, Eisen, Dale and Fennema-Notestine2010; Peper et al., Reference Peper, Brouwer, Boomsma, Kahn and Hulshoff Pol2007; Thompson et al., Reference Thompson, Cannon, Narr, van Erp, Poutanen, Huttunen, Lönnqvist, Standertskjöld-Nordenstam, Kaprio, Khaledy, Dail, Zoumalan and Toga2001). However, very few genetic variants have been identified that reliably explain a significant proportion of brain variation in human populations. Several researchers advocate the use of brain measures to empower the search for disease risk genes (Bis et al., Reference Bis, DeCarli, Smith, van der Lijn, Crivello, Fornage and . . . Seshadri2012; Meyer-Lindenberg & Weinberger, Reference Meyer-Lindenberg and Weinberger2006; Stein et al., Reference Stein, Medland, Arias Vasquez, Hibar, Senstad, Winkler and Thompson2012). If specific genetic variants can be related to brain differences, they may offer new molecular targets for drug development, and a deeper understanding of disease susceptibility and treatment response. For example, young adult carriers of a recently discovered risk allele for Alzheimer's disease (AD), CLU-C, have detectable differences in brain integrity approximately 50 years before the typical age of AD onset (Braskie et al., Reference Braskie, Jahanshad, Stein, Barysheva, McMahon, de Zubicaray, Martin, Wright, Ringman, Toga and Thompson2011), as do carriers of a common variant in an iron-overload gene, HFE (Jahanshad et al., Reference Jahanshad, Kohannim, Hibar, Stein, McMahon, de Zubicaray, Medland, Montgomery, Whitfield, Martin, Wright, Toga and Thompson2012). Brain measures related to genetic liability can be helpful for studying factors that may avert or promote disease (Gogtay et al., in press), and for defining the biological spectrum underlying psychiatric disorders (Akil et al., Reference Akil, Brenner, Kandel, Kendler, King, Scolnick, Watson and Zoghbi2010). For instance, carriers of the common Val66Met polymorphism in the brain-derived neurotrophic factor (BDNF) gene are at heightened risk for bipolar disorder (Fernandes et al., Reference Fernandes, Gama, Cereser, Yatham, Fries, Colpo, Lucena, Kunz, Gomes and Kapczinski2011) and schizophrenia (Green et al., Reference Green, Matheson, Shepherd, Weickert and Carr2011). They show detectable differences in white matter microstructure (Chiang et al., Reference Chiang, Barysheva, Toga, Medland, Hansell, James, McMahon, de Zubicaray, Martin, Wright and Thompson2010), hippocampal volume (Hajek et al., Reference Hajek, Kopecek and Hoschl2012), and prefrontal cortex morphology (Pezawas et al., Reference Pezawas, Verchinski, Mattay, Callicott, Kolachana, Straub, Egan, Meyer-Lindenberg and Weinberger2004).
The success of these candidate gene studies has led to increased interest in using neuroimaging endophenotypes to study genetic determinants of brain disorders (Hibar et al., Reference Hibar, Kohannim, Stein, Chiang and Thompson2011a; Thompson et al., Reference Thompson, Martin and Wright2010). Recently, there has been increased interest in an AD risk gene, growth factor receptor bound protein 2-associated protein or GAB2. The GAB2 gene is well characterized as a risk gene for the development of late-onset AD (Chapuis et al., Reference Chapuis, Hannequin, Pasquier, Bentham, Brice, Leber, Frebourg, Deleuze, Cousin, Thaker, Amouyel, Mann, Lendon, Campion and Lambert2008; Lin et al., Reference Lin, Tang, Han, Guo, Lin and Ma2010; Ramirez-Lorca et al., Reference Ramirez-Lorca, Boada, Saez, Hernandez, Mauleon, Rosende-Roca, Martinez-Lage, Gutierrez, Real, Lopez-Arrieta, Gayan, Antunez, Gonzalez-Perez, Tarraga and Ruiz2009; Reiman et al., Reference Reiman, Webster, Myers, Hardy, Dunckley, Zismann, Joshipura, Pearson, Hu-Lince, Huentelman, Craig, Coon, Liang, Herbert, Beach, Rohrer, Zhao, Leung, Bryden, Marlowe, Kaleem, Mastroeni, Grover, Heward, Ravid, Rogers, Hutton, Melquist, Petersen, Alexander, Caselli, Kukull, Papassotiropoulos and Stephan2007). Large meta-analyses confirm that it has a moderate effect on disease risk with an odds ratio of 1.27–1.51, using a 95% confidence interval (Ikram et al., Reference Ikram, Liu, Oostra, Hofman, van Duijn and Breteler2009; Schjeide et al., Reference Schjeide, Hooli, Parkinson, Hogan, DiVito, Mullin, Blacker, Tanzi and Bertram2009). There is convincing evidence that GAB2 is expressed throughout the brain and throughout life (Reiman et al., Reference Reiman, Webster, Myers, Hardy, Dunckley, Zismann, Joshipura, Pearson, Hu-Lince, Huentelman, Craig, Coon, Liang, Herbert, Beach, Rohrer, Zhao, Leung, Bryden, Marlowe, Kaleem, Mastroeni, Grover, Heward, Ravid, Rogers, Hutton, Melquist, Petersen, Alexander, Caselli, Kukull, Papassotiropoulos and Stephan2007; Trollmann et al., Reference Trollmann, Rehrauer, Schneider, Krischke, Huemmler, Keller, Rascher and Gassmann2010). In our previous study of 731 elderly subjects from the Alzheimer's Disease Neuroimaging Initiative (ADNI), we identified the GAB2 gene as significantly associated with morphometric brain volume differences using gene-based tests (Hibar et al., Reference Hibar, Stein, Kohannim, Jahanshad, Saykin, Shen, Kim, Pankratz, Foroud, Huentelman, Potkin, Jack, Weiner, Toga and Thompson2011b). We showed also that gene-based tests — based on principal component regression (PCReg) encoding the set of single nucleotide polymorphisms (SNPs) in a gene — have more power to detect significant associations than standard univariate statistical methods, in certain cases. The association of GAB2 with morphometric brain volume differences was undetectable with traditional univariate methods, but showed significant effects in the brain when the cumulative evidence of variation across the gene was incorporated into a gene-based association statistic. Another prior analysis of GAB2 in a separate, independent dataset found that a protective GAB2 haplotypic variant is associated with increased glucose metabolism in brain regions typically affected in AD (including the left temporal lobe, right frontal lobe, bilateral parietal lobes, and the precuneus) measured with fluorodeoxyglucose positron emission tomography (FDG-PET) (Liang et al., Reference Liang, Chen, Lee, Sidhar, Corneveaux, Allen, Myers, Villa, Meechoovet, Pruzin, Bandy, Fleisher, Langbaum, Huentelman, Jensen, Dunckley, Caselli, Kaib and Reiman2011). In addition, Liang et al. (Reference Liang, Chen, Lee, Sidhar, Corneveaux, Allen, Myers, Villa, Meechoovet, Pruzin, Bandy, Fleisher, Langbaum, Huentelman, Jensen, Dunckley, Caselli, Kaib and Reiman2011) showed, when studying APOE4 allele carriers specifically, that carriers have increased glucose metabolism if they also carry the protective GAB2 haplotype. Lower levels of glucose metabolism in the brain tissue of AD and mild cognitive impairment (MCI) patients may reflect a decrease in neuronal density, as well as abnormal metabolism (Magistretti & Pellerin, Reference Magistretti and Pellerin1996; Mark et al., Reference Mark, Pang, Geddes, Uchida and Mattson1997; Piert et al., Reference Piert, Koeppe, Giordani, Berent and Kuhl1996). While the Liang et al. (Reference Liang, Chen, Lee, Sidhar, Corneveaux, Allen, Myers, Villa, Meechoovet, Pruzin, Bandy, Fleisher, Langbaum, Huentelman, Jensen, Dunckley, Caselli, Kaib and Reiman2011) study focused on haplotype blocks, certain gene-based tests, such as PCReg performed by Hibar et al., (Reference Hibar, Stein, Kohannim, Jahanshad, Saykin, Shen, Kim, Pankratz, Foroud, Huentelman, Potkin, Jack, Weiner, Toga and Thompson2011b), may be considered comparable, as the GAB2 gene lies in a single linkage-disequilibrium (LD) block (Reiman et al., Reference Reiman, Webster, Myers, Hardy, Dunckley, Zismann, Joshipura, Pearson, Hu-Lince, Huentelman, Craig, Coon, Liang, Herbert, Beach, Rohrer, Zhao, Leung, Bryden, Marlowe, Kaleem, Mastroeni, Grover, Heward, Ravid, Rogers, Hutton, Melquist, Petersen, Alexander, Caselli, Kukull, Papassotiropoulos and Stephan2007). This replication of GAB2 effects in a separate sample and imaging modality lends credibility to GAB2 as a risk gene that influences the brain. It is then reasonable to suspect that GAB2 gene variants have significant, observable effects on brain morphology, perhaps even in early adulthood. However, no studies have examined how GAB2 relates to brain structure in young adults, long before the onset of neurodegeneration. In this study, we hypothesized that gene-based tests in the AD-risk gene, GAB2, would reveal significant effects on brain morphology in young adults.
Methods
Subjects
A total of 755 healthy, young adult (mean = 23.1, SD = 3.1 years) twins and their siblings (469 females) from 439 families (294 dizygotic twins, 222 monozygotic twins, 143 singletons, three dizygotic trios, and 87 siblings) were examined in this study. All subjects had standard T1-weighted MRI brain scans and genome-wide genotyping information available. All subjects were of European ancestry and recruited as part of the Queensland Twin Imaging Study (QTIM) in Australia. The QTIM study is an ongoing five-year longitudinal study of healthy young twins with structural and functional MRI, diffusion tensor imaging (DTI), genetics, and comprehensive cognitive assessments (de Zubicaray et al., Reference de Zubicaray, Chiang, McMahon, Shattuck, Toga, Martin, Wright and Thompson2008). Subjects were excluded if they reported any pathologies known to affect the brain, head injuries, or major illnesses. All subjects were right-handed, as determined by the 12-item Annett's handedness questionnaire (Annett, Reference Annett1970). Informed consent was obtained from each subject and the study was approved by the institutional review boards of the University of California, Los Angeles, and the Queensland Institute of Medical Research.
Genotyping and Quality Control Filtering
Genome-wide genotype data were collected on the Human610-Quad BeadChip (Illumina, Inc., San Diego, CA). Several SNPs were dropped from the dataset based on standard quality control filtering measures used in other large GWAS analyses (Wellcome Trust Case Control Consortium, 2007). Individual SNPs were removed based on the following criteria: call rate < 95% (8,447 SNPs removed), minor allele frequency < .01 (33,347 SNPs removed), significant deviation from Hardy-Weinberg equilibrium p < 1 × 10−6 (2,841 SNPs removed), autosomal chromosomes only, and a platform-specific score of .07 to eliminate blank genotype calls (results in a variable number of missing genotypes for each SNP).
After filtering, the hard genotype calls were imputed to the HapMap Phase II release 21 reference dataset (Altshuler et al., Reference Altshuler, Gibbs, Peltonen, Dermitzakis, Schaffner and Yu2010). Imputation was performed using the robust and freely available software, MaCH (Abecasis et al., Reference Abecasis, Li, Willer, Ding and Scheet2010). The imputed genotypes were further filtered based on minor allele frequency < .01 (38,481 SNPs removed) and R 2 < .3 (54,337 SNPs removed). After imputation, 2,439,807 SNPs passed all quality control criteria.
In this study, we were only interested in SNPs found in the candidate gene, GAB2, as it was strongly implicated in our prior study of a different dataset (Hibar et al., Reference Hibar, Stein, Kohannim, Jahanshad, Saykin, Shen, Kim, Pankratz, Foroud, Huentelman, Potkin, Jack, Weiner, Toga and Thompson2011b). We used the gene annotation function in the KGG software package (Li et al., Reference Li, Gui, Kwan and Sham2011; Li et al., Reference Li, Sham, Cherny and Song2010) to select SNPs from our set of imputed genotypes in the GAB2 gene group. SNPs within 50 Kb upstream or downstream of the GAB2 gene border were included in the final group of selected SNPs.
Image Acquisition and Processing
High resolution structural MRI scans were obtained for each subject on a single 4-Tesla scanner (Bruker Medspec). T1-weighted images were acquired with an inversion recovery rapid gradient echo sequence (TI/TR/TE = 700/1500/3.35 ms; flip angle = 8°; slice thickness = 0.9 mm, with a 2562 acquisition matrix).
Non-brain regions were removed from the T1-weighted scans using Robex (Iglesias et al., Reference Iglesias, Liu, Thompson and Tu2011) specifically trained on manually edited skull-stripped images. Next, scans were corrected for image field non-uniformity using FreeSurfer (http://surfer.nmr.mgh.harvard.edu/), and linearly aligned to a common template using a 9-parameter model (Holmes et al., Reference Holmes, Hoge, Collins, Woods, Toga and Evans1998).
3D Maps of Morphometric Brain Differences
The minimum deformation template (MDT) represents a nonlinear average of anatomical differences throughout the brain, and is used as a reference template to help compare the brain structure of subjects in the study. Using the MDT as a target, 3D ‘Jacobian’ maps of regional brain volume differences were generated for each subject, with a nonlinear, inverse consistent registration algorithm (Leow et al., Reference Leow, Huang, Geng, Becker, Davis, Toga and Thompson2005), and then downsampled to a 2 × 2 × 2 mm isometric voxel size. Each voxel value in the 3D maps measures the difference in volume between the subject's image and the template, based on the gradient of the deformation field required to deform the subject's scan to match the common template. As subjects are all registered to the same MDT, we can examine regional morphometric differences in brain volume by analyzing the determinant of the Jacobian matrix at the same voxel in each subject. In this study, we performed a whole-brain analysis of the volume differences from these 3D morphometric maps for each voxel in the brain and across all subjects.
Gene-Based Tests
Tests of association of the GAB2 gene with whole-brain, voxel-wise regional brain volume differences were conducted inside the whole-brain mask of the MDT, using PCReg (Wang & Abbott, Reference Wang and Abbott2008). PCReg test statistics are generated by first performing principal components analysis (PCA) on a set of n SNPs across our sample of subjects. The PCA outputs a set of orthogonal eigenvectors that represent the variance components from the set of SNPs. This transformation is defined so that the first principal component has the largest possible variance (accounts for as much as possible of the variability in the data), and each succeeding component in turn accounts for the highest variance possible under the constraint that it be orthogonal to (linearly uncorrelated with) the preceding components. Eigenvectors are then selected (in descending order of the amount of variance explained) until they explain at least 95% of the total variance in the original SNP set. These PCA steps provide an efficient means for dimension reduction but, more importantly, they provide a means to test the cumulative evidence for association across a full gene without being vulnerable to problems due to multicollinearity. Association tests with PCReg use a multiple partial-F test framework, which works by fitting two separate regression models (‘full’ and ‘reduced’) such that the full model contains a set of eigenvectors with age and sex covariates as regressors, and morphometric volume values as the dependent variable. The variance explained by the full model is then compared to the variance explained by a reduced model that fits the age and sex covariates as regressors, with the same morphometric volume values as the dependent variable. An F statistic and p-value are assigned to each voxel, based on the amount of additional variance explained by the full model compared to the reduced model. Our PCReg tests were implemented using the Efficient Mixed-Model Association (EMMA) software package (Kang et al., Reference Kang, Zaitlen, Wade, Kirby, Heckerman, Daly and Eskin2008). EMMA uses a kinship matrix to control for relatedness in family-based and twin samples, such as this one, using a mixed-effects model.
Multiple Comparisons Correction
Performing a large number of statistical tests at multiple voxels across the brain increases the potential of identifying false-positive findings (i.e., inflating Type I errors). To correct for the total of number of tests performed, and appropriately control Type I errors, we employed the searchlight false discovery rate procedure (searchlight-FDR; Langers et al., Reference Langers, Jansen and Backes2007). Searchlight-FDR controls for the regional significance of test statistics under the null hypothesis, and incorporates information on the spatial extent of the statistical effects and the smoothness of the underlying image. The searchlight-FDR procedure implemented in this study thresholds statistical maps at the standard q = .05 false-positive rate. Correction for the number of ‘eigen-SNPs’ included in the model is not required, as gene-based tests with PCReg evaluate association with a multiple partial-F test that includes the full set of eigen-SNPs in a single statistical test.
Results
The GAB2 Gene and PCReg
After all quality control filtering, imputation, and annotation, there were 51 SNPs in the GAB2 gene grouping. Positional and functional annotation data for each SNP in the analysis are shown in Table 1. After applying PCA to the set of 51 SNPs, we found that the first 10 principal components were sufficient to explain 95% of the total variance in the full SNP set. We used this set of 10 eigen-SNPs in the PCReg association tests at each voxel across the full brain.
TABLE 1 Gene Annotation Results for the GAB2 Gene using KGG Softwarea

SNP = single nucleotide polymorphism. The starting position of GAB2 (Entrez Gene ID: 9846) on chromosome 11 is 77916336 (base pairs) with a total length of 222532 (base pairs). For each of the 51 SNPs in our analysis, we give the positional information in base pairs, annotation of possible SNP function, and a conservation score from the UCSC Genome Browser (http://hgdownload.cse.ucsc.edu/).
a see Li et al. (Reference Li, Gui, Kwan and Sham2011) & Li et al. (Reference Li, Sham, Cherny and Song2010)
Statistical Parametric Maps
The significant regions of the searchlight-FDR adjusted p-map for the whole-brain GAB2 association tests is shown in Figure 1. In total, seven clusters survived the searchlight-FDR correction (here we use cluster to mean sets of contiguous voxels, even though we used a voxel-wise correction method). The associated regions after correction for multiple comparisons are summarized in Table 2.
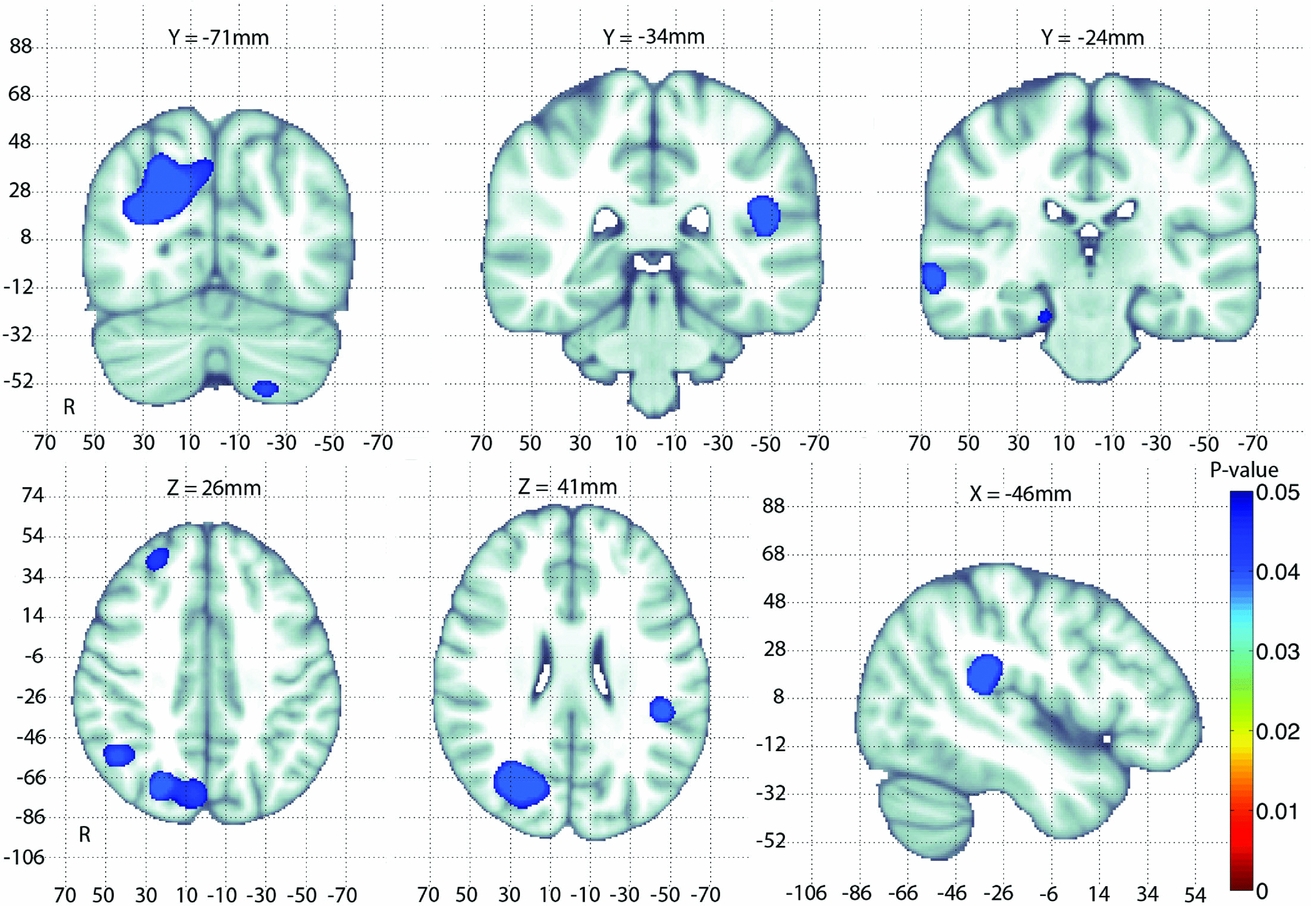
FIGURE 1 An adjusted p-map of GAB2 association tests after correcting for multiple comparisons using searchlight FDR. Adjusted p-values < .05 are considered significant and are overlaid on the MNI-152 T1 template for anatomical reference. The largest clusters of significant associations of GAB2 with morphological differences occur in the right parietal lobe.
TABLE 2 A Summary of Clusters Significantly Associated with GAB2 after Correction for Multiple Comparisons Using Searchlight FDR

MNI = Montreal Neurological Institute. Here we define ‘cluster’ to mean any set of significant contiguous voxels after correction for multiple comparisons. The location of the most highly associated voxel in each cluster is given in MNI coordinates: MNI(x,y,z). The size or extent of each cluster is given in mm3. The minimum corrected p-value is the most highly associated voxel in a given cluster in the searchlight-FDR corrected map; uncorrected p-values for that voxel are given in parentheses.
Post Hoc Analysis
The multiple partial F-test yields F-test statistics that are non-directional (in other words, they do not tell you the ‘direction’ of an effect). So, for our primary analysis, we report only p-value statistics for each voxel that passes regional FDR correction, using searchlight-FDR. These p-values only indicate that there was a significant association between GAB2 and volume differences in the cohort at a given voxel, not its direction. To get a better idea of the direction of an effect, we performed a post hoc analysis using the first principal component from the PCA of GAB2 SNPs as a regressor, along with age and sex covariates, in a mixed-effect regression (MER) model (as implemented in EMMA), at each point across the brain. In this way, we were able to obtain approximate directional effects of the GAB2 gene (i.e., those implied by the first principal component) on brain volume differences using the Beta coefficients from the MER model (Figure 2).
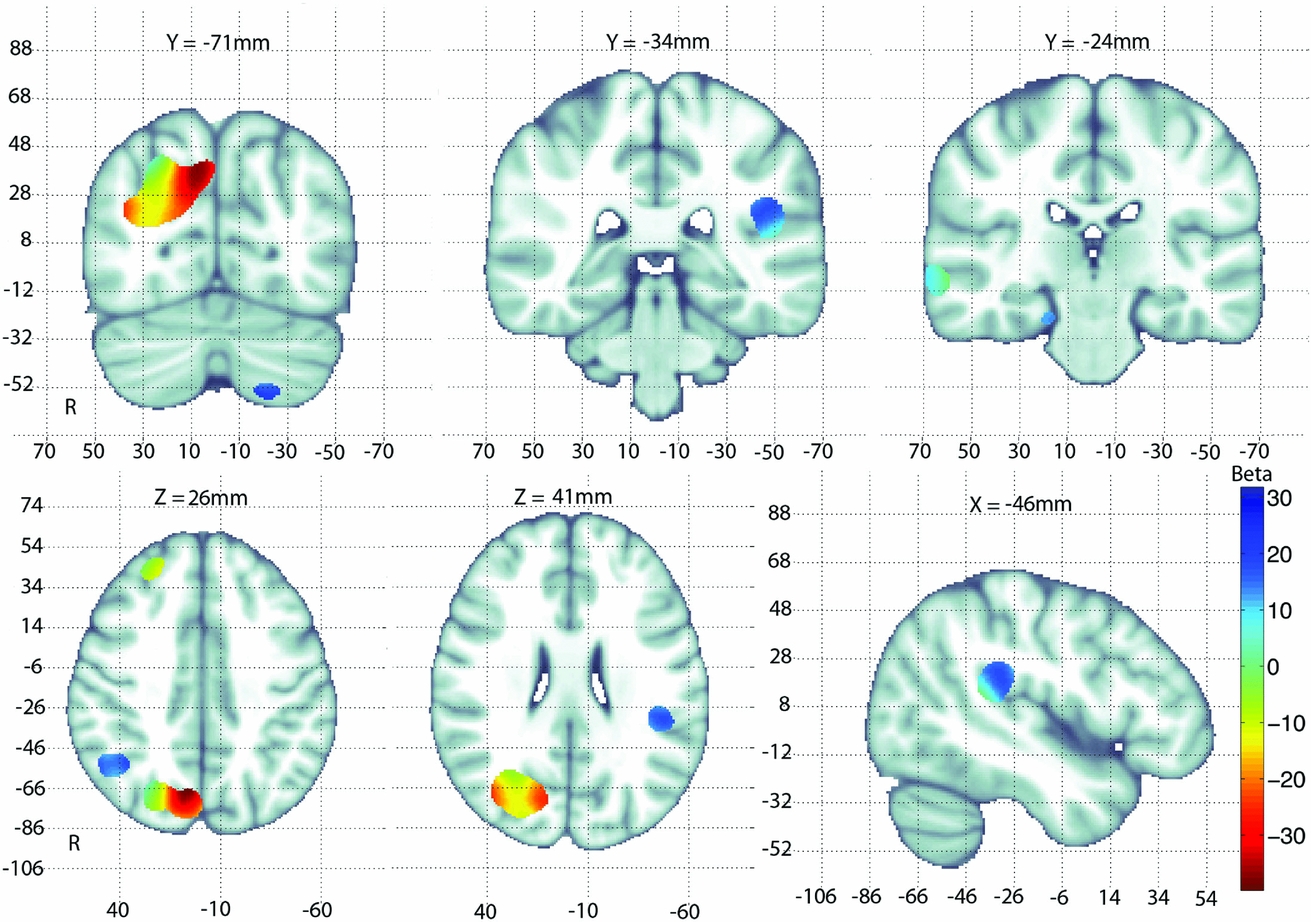
FIGURE 2 Beta coefficient values are shown for the first principal component of the GAB2 SNPs used in a multiple linear regression with age and sex as covariates. Only regions that were significant after correction for multiple comparisons using searchlight FDR are shown. The coefficient maps are overlaid on the MNI-152 T1 template for anatomical reference. Negative Beta values (warmer colors) in the tissue of the parietal lobe indicate a negative relationship of tissue volume and GAB2 loading. Positive Beta values (cooler colors) in and around the cerebrospinal fluid of the lateral sulcus and temporal lobes indicate a positive relationship of CSF expansion with GAB2 loading.
Discussion
Here we found a significant association between morphological differences in young-adult brains and variation in the GAB2 gene. These results augment our previous work with the elderly ADNI sample that demonstrated for the first time that variation in the GAB2 gene was associated with observable morphological differences in the brain. Our study of the QTIM sample expands on the literature that has established GAB2 as an AD risk gene. Prior studies have not considered how this gene might affect the brain long before the typical age of onset of the disease. We found significant differences in regional brain volume in a large region of the right parietal lobe, and additional smaller significant clusters in the left parietal, and along the tissue/cerebral spinal fluid (CSF) boundary, in the temporal lobes of the young-adult twins. A post hoc analysis was also performed to determine the direction of the GAB2 gene effects on brain morphology. There was a negative correlation between overall tissue volume and GAB2 loading in the right parietal lobe region. In addition, we found a positive correlation between GAB2 loading and CSF volume along the sulci of the temporal lobes (partial volume effects along the tissue/CSF boundary means that voxels along the border usually represent the effect of CSF volume change), as shown in Figure 2.
Function of GAB2
GAB2 encodes a human adapter protein that acts in a number of cell proliferation pathways, especially in endothelial cells (Zhang & Broxmeyer, Reference Zhang and Broxmeyer2000) and is expressed throughout the human brain (Zhao et al., Reference Zhao, Yu, Shen and Feng1999). The mechanism by which the GAB2 gene leads to increased neurodegeneration is well understood. Reiman et al. (Reference Reiman, Webster, Myers, Hardy, Dunckley, Zismann, Joshipura, Pearson, Hu-Lince, Huentelman, Craig, Coon, Liang, Herbert, Beach, Rohrer, Zhao, Leung, Bryden, Marlowe, Kaleem, Mastroeni, Grover, Heward, Ravid, Rogers, Hutton, Melquist, Petersen, Alexander, Caselli, Kukull, Papassotiropoulos and Stephan2007) showed that normally functioning GAB2 protein is responsible for suppressing the phosphorylation of tau tangles associated with the development of Alzheimer's disease. Using small-interfering RNA (siRNA) to knock-down GAB2 function, Reiman et al. also demonstrated a significant increase in tau tangle formation in neuronal tissue with the GAB2 protein function diminished. While the role of GAB2 in the pathway that leads to the formation of tau tangles is well understood, there is scant evidence in the literature about functional mechanisms and pathways through which changes in the efficiency of the GAB2 protein might lead to morphological differences in young-adult brains.
Knowing that an AD risk gene such as GAB2 may have a detectable effect in the young adult brain is useful, as it may indicate a developmental vulnerability to AD, without directly promoting AD pathology (e.g., amyloid plaque and neurofibrillary tangle formation). This is the most likely scenario as the young-adult brain is fairly low in amyloid deposits (Bartzokis, Reference Bartzokis2011; Braak & Braak, Reference Braak and Braak1997). Even so, recent studies of some other AD risk genes have also revealed detectable effects in young adults. Shaw et al. (Reference Shaw, Lerch, Pruessner, Taylor, Rose, Greenstein, Clasen, Evans, Rapoport and Giedd2007) showed that adolescent APOE4 allele carriers have a thinner cortex and slower cortical thickening than their APOE2/3 counterparts. A genetic variant (rs11136000) in the AD risk gene, CLU, found in 88% of Caucasian subjects, confers nearly a 20% lifetime increase in the risk for AD and is associated with white matter differences in DTI scans of young adults (Braskie et al., Reference Braskie, Jahanshad, Stein, Barysheva, McMahon, de Zubicaray, Martin, Wright, Ringman, Toga and Thompson2011). Similarly, the H63D variant in HFE, an iron overload gene, is associated with white matter microstructure in young adults and is thought to be associated with AD risk (Jahanshad et al., Reference Jahanshad, Kohannim, Hibar, Stein, McMahon, de Zubicaray, Medland, Montgomery, Whitfield, Martin, Wright, Toga and Thompson2012).
Conclusions and Interpretations
An essential part of endophenotype theory suggests that complex brain disorders may be better described by traits related to a disease, but with simpler genetic determinants than case-control phenotypes (Gottesman & Gould, Reference Gottesman and Gould2003). Neuroimaging endophenotypes are now widely used to assess genetic contributions to complex neurodegenerative disorders such as late-onset Alzheimer's disease (LOAD). Given the cost and difficulty of collecting large neuroimaging datasets, it can be challenging to collect enough data for unbiased gene discovery methods such as GWAS (de Geus, Reference de Geus2010). Very large neuroimaging genetics consortia are only just beginning to aggregate samples large enough to discover and replicate GWAS findings from imaging studies (ENIGMA; http://enigma.loni.ucla.edu; Stein et al., Reference Stein, Medland, Arias Vasquez, Hibar, Senstad, Winkler and Thompson2012). Even so, hypothesis-driven candidate gene studies may be performed in smaller datasets, without such a heavy correction for the number of SNPs assessed. Several such studies of disease risk genes (e.g., CLU, Braskie et al., Reference Braskie, Jahanshad, Stein, Barysheva, McMahon, de Zubicaray, Martin, Wright, Ringman, Toga and Thompson2011; and HFE, Jahanshad et al., Reference Jahanshad, Kohannim, Hibar, Stein, McMahon, de Zubicaray, Medland, Montgomery, Whitfield, Martin, Wright, Toga and Thompson2012) have revealed how variants in these genes affect the brain, offering a plausible mechanistic explanation of how they may promote risk for neurological disease. Endophenotypes can help to localize gene effects, revealing functional and anatomical differences between carriers of different variants; they may also help to classify disease subtypes that may not be apparent in case-control studies (de Geus, 2010).
The current study shows detectable associations between the AD risk gene GAB2 and morphological brain differences in young adults. Our findings build on our previous work in a separate cohort of elderly subjects that shows significant morphological differences associated with variation in the GAB2 gene (Hibar et al., Reference Hibar, Stein, Kohannim, Jahanshad, Saykin, Shen, Kim, Pankratz, Foroud, Huentelman, Potkin, Jack, Weiner, Toga and Thompson2011b). While we report significant associations in the right parietal lobe, lateral sulcus, and bilateral temporal lobes, none of the voxels overlap with the voxels that survived statistical thresholding in the original ADNI sample. In this analysis, we performed an unbiased search across each voxel of the full brain without incorporating information from our prior tests in the ADNI dataset. The reason for this is that voxels that are significant after correction for multiple comparisons do not necessarily represent the only voxels where GAB2 may have an effect. We wanted to perform an analysis that allowed for the possibility to observe an effect anywhere in the brain. As such, the most appropriate test is a brain-wide search, as it asserts only that the gene has some effect in the brain, rather than making a stronger assertion about its specific localization. In addition, the QTIM dataset differs substantially from the ADNI dataset (subjects are, on average, around 50 years younger in the QTIM study, and are scanned at a different field strength on a different continent). Given this, the regions where GAB2 is associated with brain morphology may be very different from those observed in the elderly ADNI sample. From a biological point of view, it may also be that a gene has regional effects on the brain that either spread out or become more regionally specific over the human lifespan. Or, perhaps more likely, it could be that a very weak effect is spread over the entire brain in both samples, but due to noise and anatomical variability in the two cohorts, different locations in the brain contribute to the statistical results in each cohort. As an alternative, more stringent, approach, a conjunction test (Nichols et al., Reference Nichols, Brett, Andersson, Wager and Poline2005) could be performed to directly identify voxels where a gene effect is statistically significant in several cohorts at once. Such tests have relatively low power, as they require that the gene effect be found consistently in the same voxels in all cohorts, rather than across the brain in aggregate in each cohort. Different formulations of the null hypothesis may therefore greatly affect the power and scope of the inferences. We found a large set of significant voxels in the parietal and temporal lobes that were associated with tissue volume differences. Regions of both the parietal and temporal lobes have been associated with susceptibility for and progression of Alzheimer's disease (Leow et al., Reference Leow, Yanovsky, Parikshak, Hua, Lee, Toga, Jack, Bernstein, Britson, Gunter, Ward, Borowski, Shaw, Trojanowski, Fleisher, Harvey, Kornak, Schuff, Alexander, Weiner, Thompson and Neuroimaging Initiative2009). Parietal lobe atrophy (Scahill et al., Reference Scahill, Schott, Stevens, Rossor and Fox2002), impaired white matter microstructure (Bozzali et al., Reference Bozzali, Falini, Franceschi, Cercignani, Zuffi, Scotti, Comi and Filippi2002), and glucose hypometabolism (Langbaum et al., Reference Langbaum, Chen, Lee, Reschke, Bandy, Fleisher, Alexander, Foster, Weiner, Koeppe, Jagust, Reiman and Neuroimaging Initiative2009), are well-studied effects of Alzheimer's disease progression on the brain. Additionally, Liang et al. (Reference Liang, Chen, Lee, Sidhar, Corneveaux, Allen, Myers, Villa, Meechoovet, Pruzin, Bandy, Fleisher, Langbaum, Huentelman, Jensen, Dunckley, Caselli, Kaib and Reiman2011) showed reduced glucose metabolism bilaterally in the parietal and temporal lobes of carriers of a GAB2 haplotypic risk variant. Many of the significant regions from that study appear to overlap with the regions found in this study, especially in the parietal lobes. Volume changes in the temporal lobes, specifically, have been commonly used as a biomarker for tracking the progression of Alzheimer's disease, and also for image-based diagnostic classification (Jack et al., Reference Jack, Petersen, Xu, O'Brien, Smith, Ivnik, Tangalos and Kokmen1998; Jack et al., Reference Jack, Petersen, Xu, Waring, O'Brien, Tangalos, Smith, Ivnik and Kokmen1997). Here we found that GAB2 is associated with differences in regional brain volumes in young adults, in regions strongly implicated in the progression of Alzheimer's disease. This adds support to the notion that GAB2 may be used to help identify individuals at heightened risk for AD long before the onset of disease.
Further replication of observed effects of the GAB2 gene in young adults is still required in order to aggregate evidence about true morphological effects. While searching for genetic determinants of morphological brain differences, the interpretation of results should consider not only individual genetic contributions, but potentially correlated alleles as well. In our study, we detected a very mild positive correlation of the 51 SNPs from the GAB2 gene and the CLU Alzheimer's disease risk SNP rs11136000 (Pearson's r = .07–.08), though none of the test statistics passed a Bonferroni correction for the number of SNPs tested (p < 9.8 × 10−4). Tests of other major candidate gene SNPs, such as the BDNF Val66Met polymorphism (Chiang et al., Reference Chiang, Barysheva, Toga, Medland, Hansell, James, McMahon, de Zubicaray, Martin, Wright and Thompson2010), and the H63D polymorphism of the HFE gene (Jahanshad et al., Reference Jahanshad, Kohannim, Hibar, Stein, McMahon, de Zubicaray, Medland, Montgomery, Whitfield, Martin, Wright, Toga and Thompson2012), showed no evidence of positive correlation. There could, in principle, be correlations between risk alleles, especially in an elderly population, due to a survivor effect. This could arise if adverse variants in a gene were associated with early mortality. Protective variants might then be present together in the survivors more frequently than their random co-occurrence in randomly selected people. Our analysis provides new evidence for an association between morphological differences and variation in the GAB2 in young adults, long before the onset of AD pathology. We hope that our findings will inform future research on the functional relevance of GAB2 in neural development.
Acknowledgments
We thank the twins for their participation, Kori Johnson and the radiographers for MRI scanning and preprocessing the images, Marlene Grace and Ann Eldridge for twin recruitment, Daniel Park for database support, Anjali Henders for DNA processing and preparation, Scott Gordon for quality control and management of the genotypes. The QTIM study was supported by the National Institute of Child Health and Human Development (R01 HD050735), and the National Health and Medical Research Council (NHMRC 486682,1009064), Australia. Genotyping was supported by NHMRC (389875). DPH is partially supported by NSF GRFP grant DGE-0707424. OK was supported in part by the UCLA MSTP. JLS is partially supported by a T32 post-doctoral training grant in Neurobehavioral Genetics. NJ was additionally supported by NIH NLM Grant T15 LM07356. GM was supported by an NHMRC Fellowship 613667; GZ was supported by an ARC Future Fellowship.




