


Introduction
Habitat selection is a hierarchical process of behavioural responses that may result in a disproportionate use of habitats for the survival and fitness of individuals (Johnson, Reference Johnson1980; Block & Brennan, Reference Block and Brennan1993). Studies of habitat selection have assumed a new urgency as a result of the importance of incorporating both habitat and demographic information into conservation planning (Caughley, Reference Caughley1994). An understanding of habitat selection and an explicit consideration of fitness consequences is a precondition for enhancing the survival of species (Tirpak et al., Reference Tirpak, Giuliano and Miller2008) and needs to be carefully considered in conservation planning.
Black-necked cranes Grus nigricollis live their entire life on high plateaus, primarily in western China, with the eastern population breeding on the Qinghai–Tibet Plateau and wintering on the lower Yunnan–Guizhou Plateau. The global population is estimated to be c. 8,000 (Li & Li, Reference Li and Li2005) and the species is categorized as Vulnerable on the IUCN Red List (BirdLife International, 2009) and listed in Appendix I of CITES (Inskipp & Gillett, Reference Inskipp and Gillett2005). Dashanbao National Nature Reserve in north-east Yunnan plays an important role in the conservation of the species (Kong, Reference Kong2008), with the largest population wintering on the Yunnan–Guizhou Plateau, and the Reserve is one of the most important stopover sites for the eastern black-necked crane population (Li & Yang, Reference Li and Yang2003; Qian et al., Reference Qian, Wu, Gao, Zhang, Li and Zhong2009). The species has been little studied because of the remoteness of its breeding and wintering areas (Meine & Archibald, Reference Meine and Archibald1996). Loss and degradation of habitat as a result of increasing anthropogenic pressures are the principal threats to this species, especially in its winter range (Yang, Reference Yang and Harris1991; Bishop, Reference Bishop2006).
Although there have been many studies of habitat selection by crane species (Díaz et al., Reference Díaz, González, Muňoz-Pulido and Naveso1996; Ma et al., Reference Ma, Wang and Tang1999; Végvári, Reference Végvári2002; Avilés, Reference Avilés2004) this is not the case for the black-necked crane. Of the few studies, Zhou et al. (Reference Zhou, Ding, Wang, Archibald and Pasquier1987) observed black-necked cranes selecting marshland as their main winter habitat in Caohai Reserve, Guizhou, China, and there have been descriptive analyses of the vegetation of the species’ habitat in north-east Yunnan (Li et al., Reference Li, Fang, Guan, Li, Yang and Yang2005) and of a feeding site in Dashanbao National Nature Reserve (Liu et al., Reference Liu, Kuang, Xia and Zhong2006). A study at Caohai quantified selection of winter foraging habitat by black-necked cranes, finding that sedge meadow was preferentially selected, followed by grassland and farmland (Li, Reference Li1999). These studies, however, did not include data on crane behaviour. Habitat selection is a decision-making process (Jason, Reference Jason2001) and it is important to study behaviour in habitat selection research.
Landscape changes that increase or decrease certain habitats may have significant effects on species and any such changes need to be considered by reserve managers. To improve environmental quality in central and western China the government implemented a policy in 1999 of conversion of farmland to grassland and woodland, with compensation to local farmers (Chinese State Council, 2007). Dashanbao National Nature Reserve recently implemented this policy. Based on conclusions presented in Li (Reference Li1999) we hypothesized that such land-use conversion would increase the preferred habitat available for black-necked cranes and attract more individuals to the Reserve. If the hypothesis is correct this action would be an effective conservation measure and should thus be encouraged.
In this study we focused on four aspects of habitat selection by black-necked cranes: (1) How do they use their wintering habitat? (2) What kind of habitat do they prefer (habitat preference is the consequence of habitat selection; Krausman, Reference Krausman, Launchbaugh, Sanders and Mosley1999)? (3) Which environmental factors drive habitat selection in winter? (4) What are the characteristics of habitats selected by the species?
Study area
We conducted this study between November 2006 and April 2007 in the 19,200 ha Dashanbao National Nature Reserve in north-east Yunnan Province, China. The Reserve was established in 1994 for the protection of the black-necked crane, other wintering waterbirds and the upland wetland ecosystem (Zhong & Dao, Reference Zhong, Dao, Li, Yang and Yang2005). In 2005 the Reserve was included in the Ramsar List of Wetlands of International Importance (Ramsar Convention Secretariat, 2008). The landscape of the Reserve, which lies at altitudes of 3,000–3,200 m, comprises rolling hills, plains and valleys, with rivers, streams and lakes, and patches of farmland and meadows in the valleys and on the hills. A total of c. 1,200 black-necked cranes winter in the Reserve each year, roosting at four lake-side sites (Kong, Reference Kong2008). To avoid the cranes starving during periods of heavy snow, Reserve staff provision them with corn Zea mays on the marsh near to the biggest roost at Dahaizi Lake. However, only c. 3 kg of corn has ever been supplied on any one day and no more than 50 cranes were ever attracted to eat it (KD-J, unpubl. data), indicating that artificial feeding had little impact on habitat use and selection by the cranes. Our study area covered 1,659 ha (Fig. 1) and encompassed the biggest roost, at Dahaizi Lake.

Fig. 1 Habitat distribution interpreted from a Landsat ETM+ image of the study area and transect routes in Dashanbao National Nature Reserve. The rectangle on the inset indicates the location of the main figure in north-east Yunnan, China.
Methods
Wintering cranes are gregarious and fly each morning from a communal roost to feed in surrounding areas, returning at night (Cramp & Simmons, Reference Cramp and Simmons1985). We spent 2 days every week for 22 weeks observing the cranes whilst they fed. We selected two transect routes in the study area in October 2006, before our study started, without prior knowledge of areas favoured by the species. All black-necked cranes in the study area could be detected from the 9.5 km of transects, which were along a mountain ridge (Fig. 1). The positions of all flocks were located on 1 : 50,000 maps to facilitate calculation of distances. We walked transects once per day and switched direction of travel on subsequent days. We defined flocks as being discrete if they were more than 200 m apart (Li & Ma, Reference Li and Ma1992), and every flock was considered an independent sample.
For each flock we used instantaneous scan sampling with 3-minute intervals to collect behavioural data. Five scans were made of every flock on every study day (Altmann, Reference Altmann1974). We observed another flock once the five scans were completed, and ceased scanning when cranes flew away. We categorized crane behaviours into foraging, vigilance, walking and preening. Foraging included all behaviours related to feeding, such as searching, dealing with food and swallowing. Vigilance included gazing or observing with neck stretched, ritualistic threatening and fighting. Walking included all movements except for food searching, flying and walking while in vigilance. All activities related to self-maintenance were defined as preening.
We calculated the area of the study site by analysing remotely sensed images (Landsat ETM+. path/row 129/041, resolution 15 × 15 m; June 2001). We ground-truthed habitat composition using 105 point observations 50 m apart along the transects. Five habitat categories were distinguished: farmland, grassland, marsh, water and man-made woodland (Fig. 1). Farmland included ploughed and unploughed cereal and potato fields and Rorippa indica fields. Grassland comprised meadows with minimal water. Marsh comprised both wetlands around the lake and meadows with large areas of water. Cranes usually used areas with water no deeper than 100 cm. Man-made woodland comprised areas planted with Chinese white pine Pinus armandii and Sikang pine Pinus densata.
We recorded the following details for each flock: habitat category, location, flock size, slope, slope direction (if there was slope, we regarded 1 as north and 3, 5 and 7 as east, south and west, respectively), distance to roost, distance to the nearest water, water depth and grass height. On 2 days per month we counted the number of people passing by in 30 minutes at each location where cranes occurred and this variable was regarded as disturbance.
To calculate time budgets we determined the time allocated to each activity as the percentage of scanned individuals engaging in each activity. The mean of the five scans was treated as an independent data point and used in subsequent analysis; this can reduce potential biases introduced by scan sampling (Clutton-Brock, Reference Clutton-Brock and Clutton-Brock1977). For each array of data the Kolmogorov–Smirnov test was used to check normality. Subsequently we used parametric and non-parametric tests as appropriate (Zar, Reference Zar1999). To avoid the unit-sum constraint of habitat categories (the sum of the habitat categories equals one), habitat preference (which takes into account habitat availability) was determined using the method of compositional analysis, evaluating the relative magnitude of the log-transformed ratio of every two habitat categories instead of the absolute value (Bingham & Brennan, 2004). The calculation is dij = ln(χUi/χUj)—ln(χAi/χAj), where i and j represent the ith and jth habitat category and i ≠ j; χU and χA are habitat proportion utilized and available, respectively. When dij > 0 habitat i is used more than expected relative to habitat j, otherwise habitat i is used less than expected. The equation was used to construct a ranking matrix (Aebischer et al., Reference Aebischer, Robertson and Kenward1993). A two independent samples test was used to evaluate any differences between log-transformed observed habitat preference and availability of habitat. A one-way analysis of variance was used to test for differences in the seasonality of habitat use. For post hoc multiple comparisons of time budget between different habitat categories we used Tamhane’s T2 method because the data violated the homogeneity of variance assumption (Myers & Well, Reference Myers and Well2003).
As both biotic and abiotic factors could influence habitat selection and there could be interactions amongst these factors (Krausman, Reference Krausman, Launchbaugh, Sanders and Mosley1999) we used principal components analysis (PCA) to determine the key factors that affect habitat use (McGarigal et al., Reference McGarigal, Cushman and Stafford2000). The variables used in the PCA were distance to roost (≤ 1,000, > 1,000 and ≤ 2,000, > 2,000 m), distance to the nearest water (≤ 1,000, > 1,000 m), slope (≤ 10, > 10°), slope direction (north to east, east to south, south to west, west to north), water depth (≤ 5, > 5 cm), grass height (≤ 10, > 10 cm) and number of pedestrians (≤ 5, > 5 individuals per 30 minutes). We extracted those principal components with an eigenvalue > 1. We used SPSS v. 13.0 (SPSS Inc., Chicago, USA) for statistical analysis and considered two-tailed probabilities of 0.05 for significance.
Results
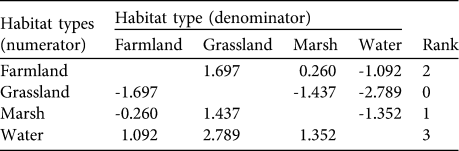
We observed 505 independent crane flocks; 55, 23, 14 and 8% of these were observed on farmland, grassland, marsh and water areas, respectively (Table 1). We did not see any cranes in man-made woodland. There was no difference in habitat utilization between months (least significant difference P > 0.05; Fig. 2). The cranes utilized the four habitats randomly (t 4 = 0.250, P = 0.815). The ranking of habitat preference based on compositional analysis was water > farmland > marsh > grassland (Table 2). There was no detectable difference in preference for the three top-ranked habitat categories (least significant difference P > 0.05) but grassland was preferred significantly less than water and farmland (largest significant difference P < 0.05).
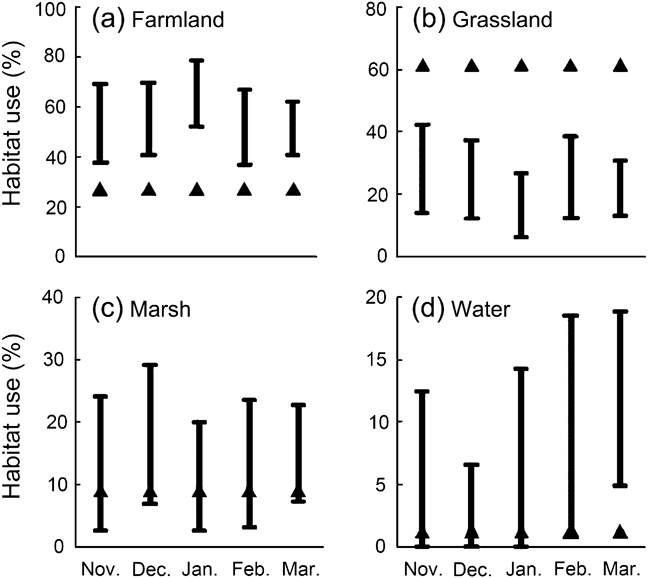
Fig. 2 Percentage use of (a) farmland, (b) grassland, (c) marsh and (d) water by black-necked cranes Grus nigricollis in the winter of 2006–2007 (November–March) in Dashanbao National Nature Reserve (Fig. 1). The black bars represent habitat use based on observations of flocks and the triangles represent expected use based on availability. Note the different y-axis dimensions.
Table 1 The area of each of the four habitats in which black-necked cranes Grus nigricollis were observed in Dashanbao National Nature Reserve (Fig. 1), with the percentage availability of each habitat, number of crane flocks observed, and percentage use by cranes of each habitat (based on observations). Man-made woodland is not included as cranes were not observed there. Although the total study area covered 1,659 ha, 154.6 ha of this is woodland and villages, and thus the total area of the four habitats is 1504.4 ha.

Table 2 The results of the evaluation of habitat preference by black-necked cranes G. nigricollis in Dashanbao National Nature Reserve (Fig. 1) using compositional analysis (see text for details). This ranking matrix was constructed with the log-transformed ratio of every two habitat category components, based on utilization and availability of habitats. Rank was determined by the number of positive values in each row (Aebischer et al., Reference Aebischer, Robertson and Kenward1993).
Time spent foraging was mostly in farmland and grassland (Fig. 3) and there was no significant difference in time spent foraging between these two habitats (Tamhane’s T2: P = 0.469). There was, however, a significant difference between these two habitats and marsh and water (Tamhane’s T2: P < 0.001). Cranes spent significantly longer preening in water than in any other habitat (Tamhane’s T2: P < 0.001) and more time preening in marsh than in farmland and grassland (Tamhane’s T2: P < 0.001). Cranes had higher vigilance in farmland compared to water (Tamhane’s T2: P = 0.026) and walking did not differ significantly among the four habitats (Tamhane’s T2: P > 0.05).

Fig. 3 Percentage of time (with 95% confidence intervals) that black-necked cranes spent in foraging, vigilance, walking or preening in farmland, grassland, marsh and water in the winter of 2006–2007 in Dashanbao Nature Reserve (Fig. 1).
The first three components of the PCA explained a total of 74.5% of cumulative variance (Table 3). PC1 (31% of variance) was strongly affected by distance to roost, grass height and water depth, and is thus related to foraging. PC2 (27.2% of variance) was strongly influenced by slope direction and degree, and could be regarded as an energy-saving related component. PC3 (16.2% of variance) was most affected by the number of passing pedestrians. In general, our result showed that > 70% of cranes used habitat close to the roost (≤ 2,000 m), with short grass (≤ 10 cm), shallow water (≤ 5cm), little disturbance (≤ 5 pedestrians passing per 30 minutes) and on slopes of < 10° oriented north to east (Table 4).
Table 3 Results of principal components analysis (components 1–3) of seven variables potentially influencing habitat selection by wintering black-necked cranes G. nigricollis in Dashanbao National Nature Reserve (Fig. 1).

Table 4 The number and cumulative frequency of the 505 independent flocks of wintering black-necked cranes observed in our defined categories of seven habitat variables in Dashanbao National Nature Reserve (Fig. 1).

Discussion
Because habitat selection refers to a series of behavioural processes (Johnson, Reference Johnson1980; Block & Brennan, Reference Block and Brennan1993) it is essential to combine time budgets together with habitat use in habitat selection analysis but this has not been done in previous studies of the black-necked crane (Li, Reference Li1999; Liu et al., Reference Liu, Kuang, Xia and Zhong2006). Compared to simply defining which kind of habitat is used most frequently by a species, our method of including behavioural data in analysis of habitat selection could help biologists and conservationists understand more fully the role of different habitats in life histories.
Black-necked cranes avoided man-made woodland, probably because of space limitations for flight as this bird prefers large open areas (Johnsgard, Reference Johsgard1983). The cranes also generally avoided grassland, preferring farmland, marsh and water; this is probably because of different levels of food availability in the four habitats, and possibly because of other physiological requirements. In Dashanbao National Nature Reserve Avena sativa, Fagopyrum tataricum, Solanum tuberosum and R. indica are the dominant crops (Zhong & Dao, Reference Zhong, Dao, Li, Yang and Yang2005), and residues are left in farmlands after harvest, providing food for the cranes. Similarly, in Tibet farmland provides a large amount of food for black-necked cranes (Bishop & Li, Reference Bishop and Li2002). A higher quantity and density of food and looser soil structure in farmland compared to grassland enhances food collection (KD-J, unpubl. data). Black-necked cranes used grassland the least in the middle of winter (especially January; Fig. 2), partly because of the frozen surface, which makes it difficult for them to find food (KD-J, pers. obs.).
We speculate that the cranes use farmland and grassland to obtain different types of food. Cereals, roots and tubers, such as A. sativa, F. tataricum, S. tuberosum and R. indica, can be found in farmland, whereas naturally occurring plants such as Pedicularis, Stellaria, Polygonatum and Veronica, and scarabid beetles, can be found in grassland (Li & Zhong, Reference Li and Zhong2010). Common cranes Grus grus in the Quercus ilex forests of Spain show a similar pattern of use of grasslands and farmlands for foraging (Díaz et al., Reference Díaz, González, Muňoz-Pulido and Naveso1996). Water, farmlands and marsh habitats provide various, but different, resources for cranes. We found that shallow water areas were used for roosting, social behaviours and as a water source (Kong et al., Reference Kong, Yang, Zhong, Dao and Zhu2008). The marsh area is important for foraging and preening activities and is also a buffer that separates the cranes from disturbance (Wu et al., 2010). A high preference of black-necked cranes for water areas, although these make up the smallest proportion of the Reserve, indicates the importance of this habitat, something also documented in a study of the black-necked crane in Napahai, Yunnan (Liu et al., Reference Liu, Yang, Yang, Zhao and Yu2010).
The PCA confirmed that a foraging related component is the main factor determining habitat selection by the black-necked crane. In Dashanbao National Nature Reserve black-necked cranes avoid places far from their roost most likely to reduce fitness and increase energy losses (Belichon et al., Reference Belichon, Clobert and Massot1996; Stamps et al., Reference Stamps, Krishnan and Reid2005), as do common cranes wintering in north-east Spain (Alonso & Alonso, Reference Alonso and Alonso1992). In habitats with short grass and shallow water the cranes may more easily be able to find food. The use of places with a shallow slope oriented north to east is probably an avoidance of the prevalent south-western winter wind, as found in a previous study in the Reserve (Liu et al., Reference Liu, Kuang, Xia and Zhong2006).
Our results suggest that black-necked cranes do not benefit from the habitat conversion taking place in Dashanbao National Nature Reserve. Conversion of farmland to woodland should be halted because the cranes never use the man-made woodland. The conversion of farmland to grassland is also not favourable to the cranes because they prefer the former. Some farmland and crop residues are necessary for the survival of black-necked cranes in the Reserve in winter. We also recommend that the Reserve protects the small area of remaining wetlands and restores more wetlands.
Acknowledgements
This study was supported by the National Key Development Program for Fundamental Research (#2007CB411600), the Western Light Associated Scholar Program of the Chinese Academy of Sciences and the International Crane Foundation Fund. We thank Dr Fan Peng-Fei for his guidance with experimental design and writing, Dr Li Feng-Shan, Dr Wu He-qi, Dr P. Buzzard and E. Doucet-Bëer for comments and suggestions, and the staff of Dashanbao National Nature Reserve for their valuable support in the field.
Biographical sketches
Kong De-Jun studies the behavioural ecology and conservation biology of the black-necked crane in China. Yang Xiao-Jun has interests in ornithological research and bird conservation. Liu Qiang works on wintering and migration ecology of the black-necked crane in China. Zhong Xing-Yao is committed to the conservation and public awareness of the black-necked crane in Dashanbao reserve and more widely in China. Yang Jun-Xing has broad interests, including animal taxonomy, evolution and biogeography of vertebrates in south-western China.







