


Introduction
Dispersal is fundamental to the genetic viability, persistence and size of populations (Harrison, Reference Harrison1991; Morris, Reference Morris1991; Hanski, Reference Hanski1999). While an animal may benefit from leaving occupied habitat to seek out vacant territory, it can also encounter higher mortality rates that typically occur during dispersal (Krebs, Reference Krebs1978; Harris & Trewhella, Reference Harris and Trewhella1988). Species distribution, and hence the potential success of reintroductions, is influenced by a species’ ability to disperse successfully (Krebs, Reference Krebs1978). Because reintroductions are, by definition, conducted where conspecifics no longer occur (IUCN, 1998), a carefully planned reintroduction will release animals in areas where resources should be relatively abundant for the first released animals. As the population increases through additional releases or population growth, the habitat may become saturated and pressures to disperse may increase. Achieving connectivity for reintroduced populations of carnivores is difficult in fragmented landscapes because home range requirements are generally large and suboptimal habitat matrices between populations can serve as high mortality sink zones (Moehrenschlager & Somers, Reference Moehrenschlager, Somers, Sillero-Zubiri, Hoffmann and Macdonald2004).
Patches of optimal short-grass prairie habitat for swift foxes Vulpes velox are disjunct throughout the northern plains as a result of extensive cultivation, and populations of swift foxes may be isolated from one another because of an inability to cross a harsh cultivated matrix (Saunders et al., Reference Saunders, Hobbs and Margules1991). Furthermore, population isolation may lead to inbreeding depression thereby affecting the long-term persistence of disjunct populations (Keller & Waller, Reference Keller and Waller2002).
The ability of swift foxes to disperse long distances is thought to be less than for larger canids (Mercure et al., Reference Mercure, Ralls, Koepfli and Wayne1993) and, in part because of limited gene flow, they have retained distinct species status from kit foxes Vulpes macrotis with which they interbreed in areas of New Mexico (Mercure et al., Reference Mercure, Ralls, Koepfli and Wayne1993). Reported dispersal distances for 48 juvenile kit foxes in California averaged only 7.8 ± SE 1.1 km (Koopman et al., Reference Koopman, Cypher and Scrivner2000) and eight juvenile swift foxes in Colorado moved just 12.6 ± SE 3.2 km from their last known den to their first discovered den after movement ceased or death occurred (Schauster et al., Reference Schauster, Gese and Kitchen2002).
Swift foxes are listed as an Endangered Species in Canada and reintroduction efforts began in 1983. Swift foxes are not listed under the Endangered Species Act in the USA but they are a Species of Concern in Montana and are of particular cultural significance to the Blackfeet Tribe in Montana, who began reintroducing swift foxes to Reservation lands in 1998. Recent studies and surveys have indicated swift fox reintroductions in both Canada and on the Blackfeet Reservation in Montana were successful (Moehrenschlager & Moehrenschlager, Reference Moehrenschlager and Moehrenschlager2001; Ausband & Foresman, Reference Ausband and Foresman2007a), with populations growing and expanding their distributions, although we believe there is still some vacant suitable habitat within both study areas. Swift fox densities near the Canadian release sites are high (Moehrenschlager & Moehrenschlager, Reference Moehrenschlager and Moehrenschlager2006) and although density estimation was not a focus of research on the Blackfeet Reservation, relative density of swift foxes near and around (≤ 20 km radius) the original release site was high (D. Ausband, unpubl. data). Genetic viability and persistence of these two populations would be enhanced through an interchange of individuals. Therefore, we wanted to assess potential dispersal of juvenile swift foxes between the two proximate but distinct reintroduced swift fox populations that collectively span the borders of Alberta and Saskatchewan in Canada and Montana in the USA. At their nearest point, foxes within these two populations are separated by mostly cropland spanning c. 200 km (Fig. 1).
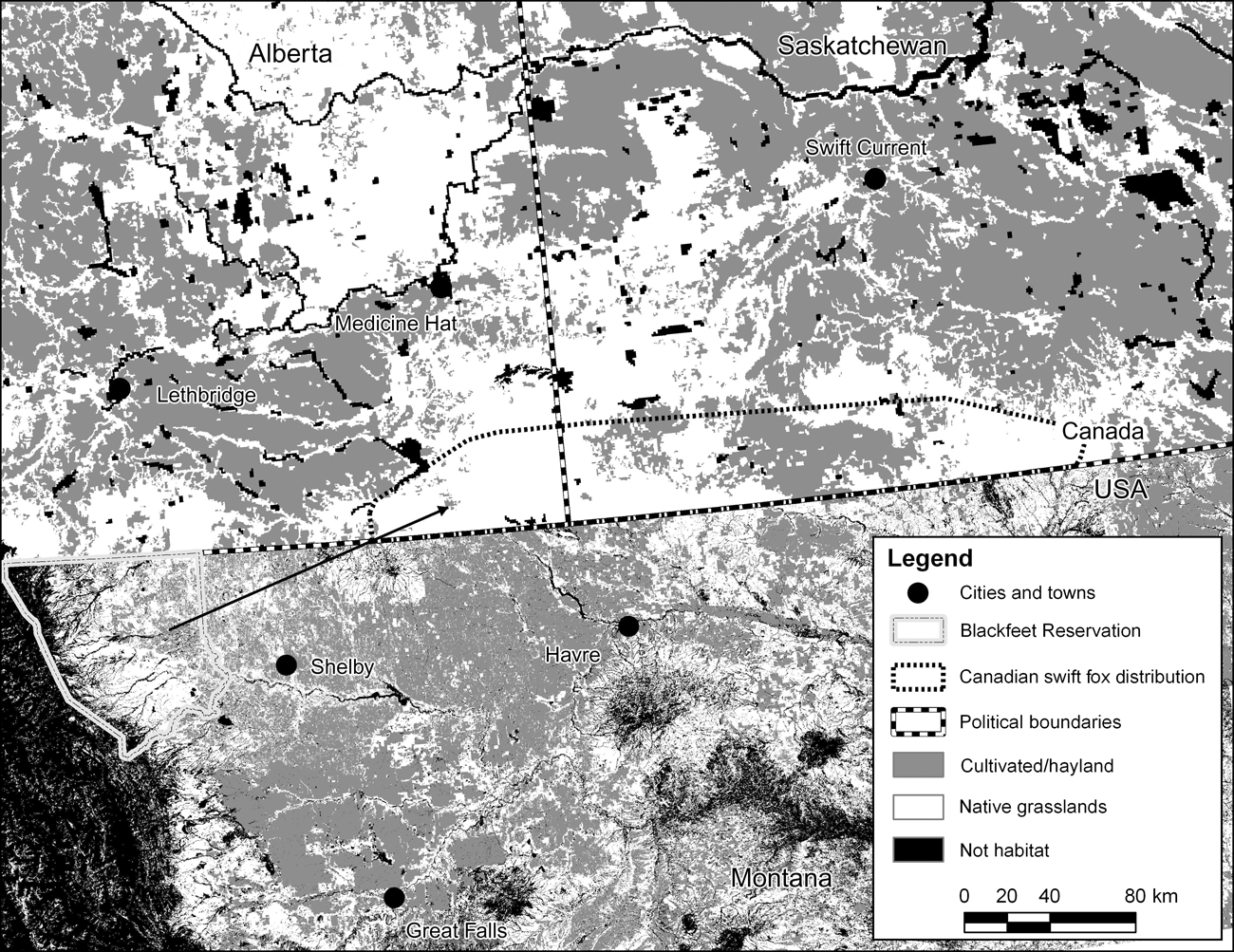
Fig. 1 Map showing grassland habitat and cropland non-habitat for swift foxes along the Canada-USA border. The straight-line arrow denotes, in simplified form, the longest movement detected of a female that dispersed from the Blackfeet Reservation, USA, to the Canadian population.
Study area
The Blackfeet Reservation, in northern Montana, USA encompasses 607,000 ha of largely short-grass prairie habitat on the eastern flank of the Rocky Mountains. Annual precipitation averages 31.8 cm and temperatures range from -40°C in January to 41°C in July. The Canadian swift fox population inhabits c. 12,000 km2 of short-grass prairie in southern Alberta and Saskatchewan. Annual temperature and precipitation are similar to that of the Blackfeet Reservation.
Methods
We live-trapped juvenile swift foxes at different times over 1995–2006 in two reintroduced populations (Fig. 1) as part of research on each individual population. Foxes were captured in box traps (Ausband & Foresman, Reference Ausband and Foresman2007a; Moehrenschlager et al., Reference Moehrenschlager, List and Macdonald2007) baited with wet cat food, sardine oil or butcher scraps. Traps were lined with wood and wire mesh to decrease the chance of injury to trapped foxes if they attempted to chew on the cage to escape (Moehrenschlager et al., Reference Moehrenschlager, Macdonald, Moehrenschlager, Carbyn and Sovada2003). Foxes caught on the Blackfeet Reservation were fitted with 48 g high frequency radiocollars, including mortality sensors and antenna lengths of 15–20 cm (Advanced Telemetry Systems, Isanti, USA), injected with transponders (AVID ID Systems, Norco, USA) between their shoulder blades, and ear-tagged or tattooed depending on location of capture and year of study. Capture and handling of animals in Montana was approved under the IACUC protocol number 036-04KFDBS-043005, and Canadian captures were approved by individual permits from Environment Canada, Alberta Fish & Wildlife, and Saskatchewan Environment and Resource Management.
During 1995–1998 swift foxes in Canada were captured in similar box traps, manually restrained, and handled by two field workers who conducted parasite counts, canine measurements, and body condition assessments. Foxes were sexed, ear-tattooed and treated with Ivermectin (Merial Limited, Iselin, USA; 0.2 mg kg-1) to combat parasite infestation, and radio-collared. Vaccinations against canine distemper, adenovirus, parainfluenza, parvovirus, and leptospirosis were administered using Duramune (Wyeth-Ayerst, St. Davids, USA; 10 mg ml-1), and Imrab (Wyeth-Ayerst, St. Davids, USA; 10 mg ml-1) provided vaccination against rabies. Radio-collars weighed 48 g, had a mortality sensor and antenna lengths of 15–20 cm. In 2000–2001 and 2005–2006, as part of a 5-year census effort, swift foxes in Canada were again captured in box traps. Canadian traps were lined completely with hardboard that was custom made with small holes; the floor, ceiling, and sides were lined with hardboard while the door and back were lined with plexiglass so that captured foxes could be seen easily (Cotterill, Reference Cotterill1997; Moehrenschlager & Moehrenschlager, Reference Moehrenschlager and Moehrenschlager2001).
Catch-and-release trapping was conducted at night (the fox activity period) to avoid heat-stress and to prevent disturbance by people. Traps were generally set at 18.00–20.00, checked at 24.00–02.00, and closed following a second check at 06.00–08.00. Trapping was not conducted at temperatures colder than –20oC or when snow, rain or wind conditions were potentially hazardous to captured foxes (Moehrenschlager & Moehrenschlager, Reference Moehrenschlager and Moehrenschlager2001). Foxes were coaxed into a denim handling bag that was placed over the end of the box trap. Every team alternated between four handling bags, which were washed frequently to reduce the transmission of fleas between foxes. Foxes were handled by both field workers of respective trapping teams. The first team member positioned the animal on his/her lap to shelter it from the wind, one hand restrained the head and covered the eyes, and the second hand restrained the body. The second field worker sexed and aged the fox, conducted parasite counts, scored body condition through palpation on a standardized index ranging from 1 (poor) to 5 (excellent), collected a hair sample for genetic analyses, checked for injuries, and tattooed the ear for identification. Foxes were uniquely marked with tattoo dye so that recaptured individuals could be easily identified.
We calculated dispersal as the straight-line distance moved between the location where a juvenile was captured in its natal range to where it was located when exploratory movement ceased or death occurred. Animals that remained within 2 km of their natal range were not considered dispersers. Some juveniles exhibited two bouts of dispersal (Ausband & Foresman, Reference Ausband and Foresman2007b), ceasing exploratory movement for ≥ 30 days only to resume exploratory movements again. We summed these straight-line distances to provide a total distance dispersed.
Results
Over 1995–1998 43 juvenile swift foxes were captured and radiocollared in the Canadian reintroduced swift fox population. In the Canadian transboundary 5-year census we captured 149 and 196 swift foxes in 2000–2001 and 2005–2006, respectively, in Canada and contiguous areas of Montana. None of these foxes were recovered on the Blackfeet Reservation and males and females dispersed relatively short distances, 12.4 ± SE 4.3 km and 10.6 ± SE 4.7 km respectively, from natal dens. Over 2003–2005 36 juvenile foxes were radiocollared and ear-tagged or pit-tagged on the Blackfeet Reservation and one of these was recovered during trapping efforts in south-eastern Alberta on 6 November 2005.
On the Blackfeet Reservation five radiocollared wild-born juveniles, three males and two females, moved relatively long total distances during 2003–2005. We report only long distance dispersal movements (> 50 km total) here because general facets of dispersal for this population are presented in Ausband & Foresman (Reference Ausband and Foresman2007b). Minimum distance dispersed averaged 50.5 ± SE 3.6 km for four of the juveniles. A fifth juvenile female fox travelled much further (191 km) and, 3 years after initial capture on the Blackfeet Reservation, was live-trapped as part of a monitoring effort in the Canadian population (Table 1). This fox had shed its radiocollar shortly after capture (> 7 days) and although we lost contact with her on the Blackfeet Reservation we were able to identify her from ear tags when she was captured in Canada. This female was caught 1 km and 3 km away from swift fox males in February 2006, 1 month before the breeding season.
Table 1 Straight-line distance moved by five wild-born, juvenile swift foxes on the Blackfeet Indian Reservation, Montana, USA, during 2003–2005. Foxes born in April/May were considered juveniles from 1 September to 1 June of the year following birth.

Discussion
Wild-born juvenile swift foxes appear capable of relatively long bouts of dispersal, and further than previously thought. In one instance a female swift fox was able to traverse the long distance between the two reintroduced populations, potentially providing genetic interchange between these seemingly disjunct populations.
With the exception of a juvenile kit fox that moved 64 km in Nevada (O'Neal et al., Reference O'Neal, Flinders, Clary and Genoways1987), the long-distance dispersal events recorded here exceed natal kit or swift fox dispersal movements recorded to date (Moehrenschlager et al., Reference Moehrenschlager, Cypher, Ralls, Sovada, List, Macdonald and Sillero-Zubiri2004; Ausband & Foresman, Reference Ausband and Foresman2007b). The lack of reported long-distance dispersal movements in contiguous swift or kit fox populations may be due to a relative abundance of prey or mates. The reintroduced northern populations, which are on the periphery of the species’ historic range, may be more resource limited; reintroduced swift foxes in Canada have larger home range requirements than conspecifics in southern populations (Moehrenschlager et al., Reference Moehrenschlager, List and Macdonald2007). Moreover, the amount of suitable habitat for reintroduced populations may be limited and foxes may be more likely to disperse over large distances as populations become saturated, although Trewhella et al. (Reference Trewhella, Harris and McAllister1988) found the opposite to be true for a large sample of urban red foxes Vulpes vulpes in Bristol, UK. Swift fox releases have shown that survival and reproduction decrease with risky long-range movements (Moehrenschlager & Macdonald, Reference Moehrenschlager and Macdonald2003) and it would seem logical that the further an animal disperses from its core range the less likely it will be to find a mate. However, successful dispersers may enhance the ability of the USA and Canadian populations to be genetically viable over time. Even a small amount of gene flow between two populations can enhance heterozygosity of individuals (Mills & Allendorf, Reference Mills and Allendorf1996), although the actual number of real migrants required may be more than one-per-generation (Wang, Reference Wang2004)
Historically, swift fox populations reintroduced in Canada and in Montana have been managed as separate entities. Our evidence of potential long-distance dispersal movements illustrates that collective recovery of these populations is possible in a metapopulation context. While movement of a swift fox was recorded from the Blackfeet Reservation into Canada we did not record movement in the opposite direction. Movement south from Canada could have occurred but went undetected because the research on the Blackfeet Reservation did not focus on the northern portion of the Reservation that borders Canada.
This research shows that swift foxes are not necessarily poor dispersers and that patches of short-grass prairie, previously thought too isolated for natural movement or colonization, should be conserved to enhance swift fox population connectivity and range. Specifically, entities currently reintroducing or recovering swift foxes (most western plains states in the USA, and Alberta and Saskatchewan, Canada) should not dismiss patches of short-grass prairie habitat that are > 25 km from the initial release site or recovery core. These seemingly isolated areas can be colonized by dispersing swift foxes and post-release monitoring programmes should also include survey efforts in these disjunct patches.
Acknowledgements
Field work in Canada was funded by Alberta Conservation Association, Alberta Fish & Wildlife, Calgary Zoo, Canadian Wildlife Service, Fairmont Palliser hotel, Husky Energy, Montana Fish Wildlife & Parks, Parks Canada, Prairie Farm Rehabilitation Agency, Saskatchewan Environment and Resource Management, Wildlife Preservation Canada, and WWF–US. Field work in the Blackfeet Reservation was funded through Defenders of Wildlife, Sigma Xi, Blackfeet Tribe and SWG. Special thanks to Cynthia Moehrenschlager, Ryan Rauscher, Pat Fargey, Brian Giddings, Elizabeth Ausband, Minette Johnson, Dan Carney, Adrian Costel and field crews who worked long hours to collect data. We appreciate comments from two anonymous reviewers that helped strengthen this article.
Biographical sketches
David E. Ausband has been participating in canid conservation in the Northern Rockies, USA, for the last 8 years. He is particularly interested in developing novel methods for long-term monitoring of wide-ranging carnivore populations. Axel Moehrenschlager is a member of the IUCN Canid and Reintroduction Specialist Groups. His Centre for Conservation Research at the Calgary Zoo conducts fundamentally significant and directly applicable research regarding reintroductions of threatened mammals, birds and amphibians.


