


Introduction
The puma Puma concolor is a generalist predator throughout its wide distribution in the Western Hemisphere (Wilson & Mittermeier, Reference Wilson and Mittermeier2009). Its diet can be affected by the abundance and vulnerability of its prey, habitat characteristics, competition with other carnivores (Currier, Reference Currier1983; Iriarte et al., Reference Iriarte, Franklin, Johnson and Redford1990b), fluctuations in its native prey populations (Novaro et al., Reference Novaro, Funes and Walker2000; Polisar et al., Reference Polisar, Maxit, Scognamillo, Farrell, Sunquist and Einseberg2003) and anthropogenic factors (Fuller & Sievert, Reference Fuller, Sievert, Gittleman, Funk, Macdonald and Wayne2001). The varied diet of the puma across its range reflects the diverse habitats it inhabits (Sunquist & Sunquist, Reference Sunquist and Sunquist2002). The puma can satisfy its food requirements by predating on either native or introduced wild and domestic species (Ackerman et al., Reference Ackerman, Lindzey and Hemker1984; Ockenfels, Reference Ockenfels1994; Wehausen, Reference Wehausen1996; Franklin et al., Reference Franklin, Johnson, Sarno and Iriarte1999; Novaro et al., Reference Novaro, Funes and Walker2000; Donadio et al., Reference Donadio, Novaro, Buskirk, Wurstten, Vitali and Monteverde2009; Foster et al., Reference Foster, Harmsen and Doncaster2010a) of a wide variety of sizes (< 1–150 kg; Iriarte et al., Reference Iriarte, Franklin, Johnson and Redford1990b). Although categorized as Least Concern on the IUCN Red List because it is a widespread species, the puma is considered to be declining and its conservation presents numerous challenges (Caso et al. 2008).
In North America the puma’s diet formerly comprised large prey, especially ungulates (Beck et al., Reference Beck, Beecham, Beier, Hofstra, Hornocker and Lindezey2005). For example, mule deer Odocoileus hemionus, pronghorn Antilocapra americana and bighorn sheep Ovis canadensis are important components of the puma’s diet in the states of Utah, Arizona and California, USA, respectively (Ackerman et al., Reference Ackerman, Lindzey and Hemker1984; Ockenfels, Reference Ockenfels1994; Wehausen, Reference Wehausen1996). In the tropical areas of Central and South America the puma’s prey includes a wide range of species and sizes, from ungulates to small mammals such as deer, and marsupials and small and large rodents (Rabinowitz & Nottingham, Reference Rabinowitz and Nottingham1986; Iriarte et al., Reference Iriarte, Franklin, Johnson and Redford1990b; Scognamillo et al., Reference Scognamillo, Maxit, Sunquist and Polisar2003; Foster et al., Reference Foster, Harmsen and Doncaster2010a). In Chilean Patagonia the diet of the puma is dominated by large-bodied native prey such as the guanaco Lama guanicoe and pudu Pudu pudu but pumas also prey on introduced species such as domestic sheep Ovis aries and European hare Lepus europaeus (Wilson, Reference Wilson1984; Yáñez et al., Reference Yáñez, Cárdenas, Gezelle and Jaksic1986; Iriarte et al., Reference Iriarte, Franklin, Johnson and Redford1990b; Franklin et al., Reference Franklin, Johnson, Sarno and Iriarte1999; Rau & Jiménez, Reference Rau and Jiménez2002).
There are few studies of the puma’s diet in Argentine Patagonia. Studies during 1984–1997 at Lihuel Calel National Park in southern La Pampa province showed that puma diet comprised mainly plains vizcacha Lagostomus maximus, a rodent weighing c. 4.5 kg (Branch et al., Reference Branch, Pessino and Villareal1996; Pessino et al., Reference Pessino, Sarasola, Wander and Besoky2001). During 1985–1994 the populations of the plains vizcacha in the park declined by > 90% (Branch et al., Reference Branch, Pessino and Villareal1996), and its occurrence as a prey item was reduced by 70% in the puma diet (Pessino et al., Reference Pessino, Sarasola, Wander and Besoky2001). Over the same period an increase in the consumption of bigger prey (the introduced wild boar Sus scrofa and red deer Cervus elaphus, and guanaco and smaller prey such as the big hairy armadillo Chaetophractus villosus and rodents) was observed (Branch et al., Reference Branch, Pessino and Villareal1996; Pessino et al., Reference Pessino, Sarasola, Wander and Besoky2001). A study in north-western Argentine Patagonia showed that most of the biomass consumed by pumas was of introduced species, particularly the European hare and the red deer, and only a small proportion was of native species, such as the big hairy armadillo and rodents (Novaro et al., Reference Novaro, Funes and Walker2000). The elevated consumption of introduced prey was the result of their higher abundance and availability compared to that of native prey (Novaro et al., Reference Novaro, Funes and Walker2000). Such an imbalance can increase until native prey do not interact significantly with their natural predators and are therefore considered ecologically extinct (Estes et al., 1989).
This concept is relevant for the conservation of extant communities because the loss of a species or its functional role may substantially affect the abundance and population structure of other species in the community. For example, guanacos may have suffered local and ecological extinctions throughout Patagonia (Novaro et al., Reference Novaro, Funes and Walker2000; Pessino et al., Reference Pessino, Sarasola, Wander and Besoky2001) because of human activities such as poaching and livestock husbandry, which has displaced the guanaco towards less favourable foraging areas (Baldi et al., Reference Baldi, Albon and Elston2001; Travaini et al., Reference Travaini, Bustamante, Rodriguez, Zapata, Procopio, Pedrana and Peck2007). Additionally, agriculture, oil exploitation and mining have altered habitats and made new areas accessible to sport hunters and poachers (Donadio & Buskirk, Reference Donadio and Buskirk2006), which could have accelerated the decline of native wildlife, both in density (Yáñez et al., Reference Yáñez, Cárdenas, Gezelle and Jaksic1986; Puig et al., Reference Puig, Videla and Cona1997) and distribution (Pedrana et al., Reference Pedrana, Rodríguez, Bustamante, Travaini and Zanón Martínez2009).
With the exception of the study of Novaro et al. (Reference Novaro, Funes and Walker2000), in Argentine Patagonia the effect of introduced species on the diet of the puma has not been described. Based on their localized observations Novaro et al. (Reference Novaro, Funes and Walker2000) concluded that the role of the guanaco and lesser rhea Pterocnemia pennata as prey for the puma had been supplanted by red deer, European hare and livestock (sheep, goats and horses), and they suggested that this may also be the case throughout this extensive and heterogeneous region.
The purpose of our study was to examine the supposed ecological extinction of the puma’s native prey in southern Patagonia and to determine if the observations of Novaro et al. (Reference Novaro, Funes and Walker2000) could be extrapolated to all of Argentine Patagonia. We analysed puma diet in three protected areas and two sheep ranches in Santa Cruz province,. As far as we are aware this is the first study to describe the puma’s diet in the largest (c. 250,000 km2) and southernmost continental province of Argentine Patagonia.
Study area
Phytogeographically the study area is in the Central Patagonian District in the Patagonian province, where the characteristic vegetation is a mixed steppe of grassland and shrubs (Cabrera, Reference Cabrera1976). The climate is dry and cold, with strong western winds (Harris, Reference Harris1998); mean annual temperature is 17 °C and mean annual precipitation varies from 125 mm in the east to 500 mm in the west. Our study was conducted in three protected areas and two sheep ranches in Santa Cruz province (Fig. 1). The protected areas, which are under the Argentine National Park Administration, were the 570 km2 Monumento Natural Bosques Petrificados, the 1,150 km2 Perito Moreno National Park and the 610 km2 Monte León National Park; the latter was created in 2000 from a sheep ranch. The sheep ranches were the 400 km2 25 de Marzo Ranch, > 100 km from the nearest protected area, the Monumento Natural Bosques Petrificados, and the 200 km2 Doraike Ranch neighbouring Monte León National Park. The Monumento Natural Bosques Petrificados and the 25 de Marzo Ranch are on the central plateau of Santa Cruz, where shrub-steppe is the dominant vegetation type. The Perito Moreno National Park is in the west of the province, where the Andean mountain range and the Patagonian steppe are the main vegetation types. The Monte León National Park and the Doraike Ranch ranch are in the south-east, on the Atlantic coast, where the dominant vegetation type is shrub-steppe.
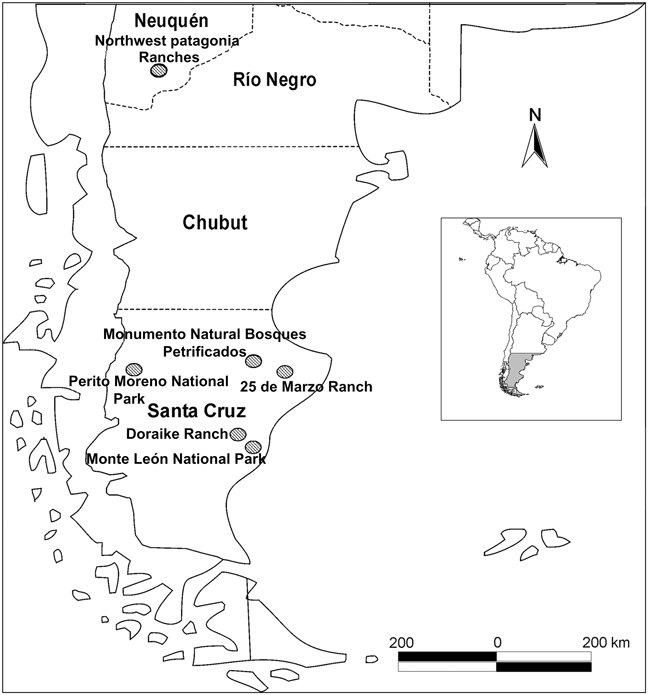
Fig. 1 Location of the five study sites in Santa Cruz province and the ranches in north-west Patagonia studied by Novaro et al. (Reference Novaro, Funes and Walker2000). The inset shows the location of the main map in South America.
Methods
Puma diet was determined by identification and quantification of prey remains in 282 scats, which were collected in the five study sites during 2004–2007. At Monumento Natural Bosques Petrificados puma scats were collected annually by searching within 4 km2 randomly selected plots whilst developing a carnivore monitoring programme based on signs (Travaini et al., Reference Travaini, Pereira, Martínez-Peck and Zapata2003). At the other four sites scats were collected specifically for this study by walking through the area, selecting the most rugged sites where puma scats are easily found.
Puma scats were identified by their size, colour and shape (Yáñez et al., Reference Yáñez, Cárdenas, Gezelle and Jaksic1986). They are easily differentiated from those of culpeo Lycalopex culpaeus and grey foxes L. griseus (Zapata et al., Reference Zapata, Travaini, Delibes and Martínez-Peck2005). The area is also inhabited by the pampas cat Leopardus colocolo and Geoffroy’s cat Leopardus geoffroyi. These scats are easily differentiated because of their smaller size and location, as the two species usually make their latrines in small caves and crooks of trees where a puma could not reach (Sunquist & Sunquist, Reference Sunquist and Sunquist2002; Zapata et al., Reference Zapata, Travaini, Delibes and Martínez-Peck2005). Collected scats were dried and stored in labelled paper envelopes until analysis.
Bones and hairs of mammalian prey were identified using a reference collection and a key of skulls and jaws (Pearson, Reference Pearson1995), or patterns of hair scales and medulla (Chehébar & Martín, Reference Chehébar and Martín1989). Bird remains, feathers, and bones were identified by comparing them with our own reference material. Identified prey items were grouped into taxa. Puma diet composition at each site was expressed as percentage occurrence (i.e. the number of times that an item appears in all scats expressed as the percentage of the total number of scats) and relative biomass of prey consumed using the equation of Ackerman et al. (Reference Ackerman, Lindzey and Hemker1984). Prey body masses were obtained from Redford & Eisenberg (Reference Redford and Eisenberg1992) and from our own records.
Results
In the five areas studied the puma preyed on at least 14 mammal and four bird species (Table 1). In the Monumento Natural Bosques Petrificados the main prey of the puma according to consumed biomass was the guanaco, followed by the European hare, the Patagonian pichi Zaedyus pichiy, and lesser rheas, although European hare was the most common prey, followed by the guanaco and other mammals (mainly rodents). Similarly, puma diet at the 25 de Marzo Ranch comprised guanaco, followed by the European hare, and another native mammal, the Patagonian mara Dolichotis patagonum.
Table 1 Diet of the puma Puma concolor in the five study sties in Argentine Patagonia (Fig. 1) during 2004–2007 as % occurrence and % consumed biomass (see text for details). Percentages of occurrences do not sum to 100 because diets are expressed as the % of total number of scats.
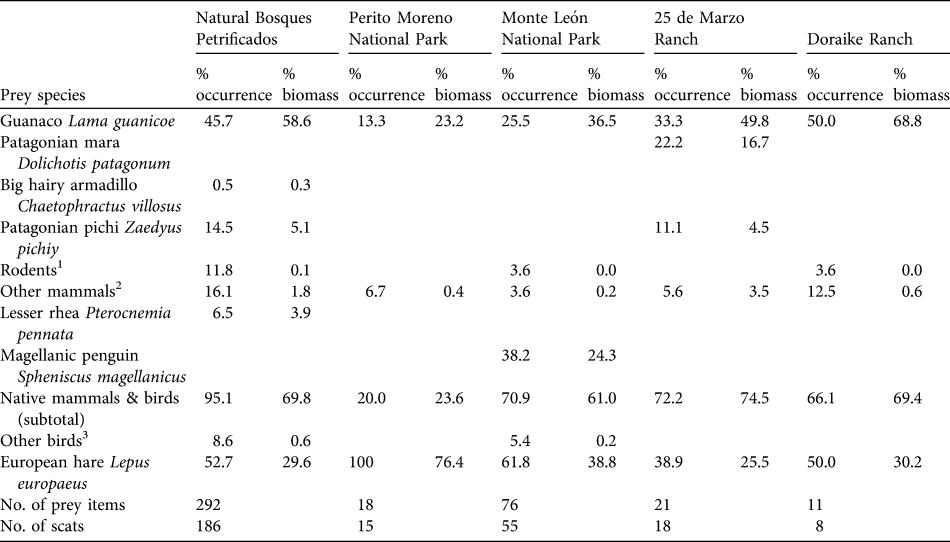
1 Includes Ctenomys magellanicus, Eligmodontia morgani, Phyllotis xanthophygus, Abrotrix xanthorinus, Graomys grisoflaveus, Microcavia australis and Reithrodon auritus.
2 Includes Mustelidae Galictis cuja and the marsupial Lestodelphys halli
3 Includes unidentified birds, Cloephaga picta and Tinamotis ingoufi
In Monte León and Perito Moreno National Parks the main prey of the puma according to consumed biomass was the European hare. The second most important prey item in these National Parks was the guanaco. The Magellanic penguin Spheniscus magellanicus, never previously described in the puma diet, was consumed in Monte León National Park. In the Doraike Ranch, which neighbours Monte León National Park, the main prey was the guanaco, followed by the European hare.
Native prey contributed > 60% of the total consumed biomass in the Monumento Natural Bosques Petrificados, Monte León National Park and their neighbouring ranches (25 de Marzo Ranch, and Doraike Ranch; Fig. 2). Native prey in Perito Moreno National Park represented only 23.6% of the consumed biomass and the introduced European hare dominated its diet, accounting for the 76.4% of the total consumed biomass (Fig. 2).
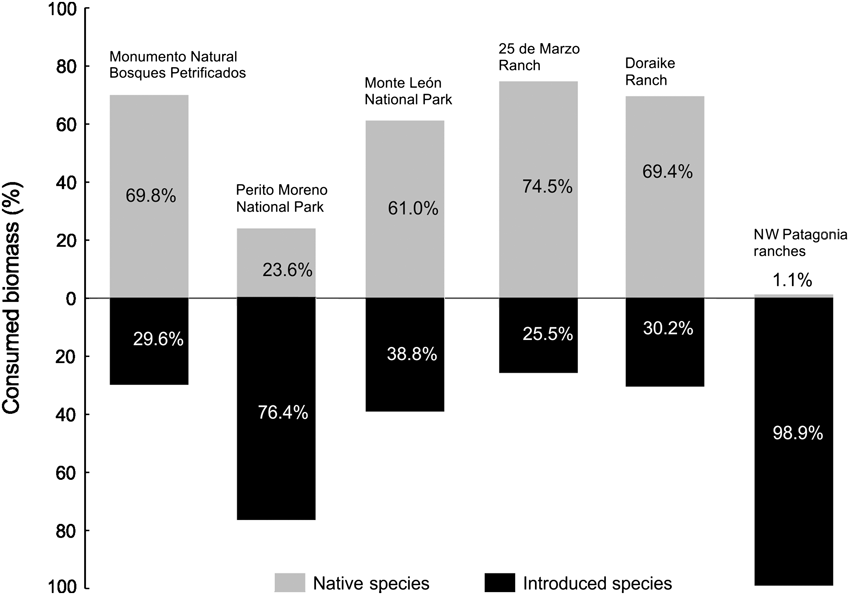
Fig. 2 Percentage of biomass of native and introduced species consumed by the puma Puma concolor at the five study sites and the ranches in north-west Patagonia, the latter from Novaro et al. (Reference Novaro, Funes and Walker2000; Fig. 1).
Discussion
In Santa Cruz province native prey species are still part of the diet of the puma. The majority of scats collected indicated that the puma’s diet there includes guanaco and variable contributions of Patagonian mara, Patagonian pichi and the Magellanic penguin. Nevertheless, the introduced European hare was also an important component and found in scats from all five sites. Since its introduction in Argentina and Chile (Grigera & Rapoport, Reference Grigera and Rapoport1983) the European hare has been heavily consumed by both terrestrial and aerial predators (Iriarte et al., Reference Iriarte, Franklin and Johnson1990a,b; Rau & Jiménez, Reference Iriarte, Franklin, Johnson and Redford2002; Zapata et al., Reference Rau and Jiménez2005). In some areas of Argentine Patagonia the European hare has become the base for the formation of new trophic guilds (Zapata et al., Reference Zapata, Travaini, Ferreras and Delibes2007) by displacing native prey from their ecological role (Lamberttucci et al., Reference Lamberttucci, Trejo, Di Martino, Sanchez-Zapata, Donazar and Hiraldo2009; Walker & Novaro, Reference Walker, Novaro, Hornocker, Negri and Rabinowitz2009). This phenomenon has been attributed to the hare’s extensive distribution, high abundance and optimal size as prey for many native predators (Donadio et al., Reference Donadio, Pauli and Bonino2005). In Perito Moreno National Park the European hare was the main component of puma scats. This protected area includes the ecotone between the Andean mountain range and the Patagonian steppe, where suitable habitat result in high densities of hares (Park Rangers, pers. comm.).
A study in Torres del Paine National Park in Chilean Patagonia found a density of 11.7 guanacos per km2 (Franklin et al., Reference Franklin, Johnson, Sarno and Iriarte1999), more than twice that found in our study area (4.7 per km2; E. Daer, National Park Administration, pers. comm.) but guanacos constituted the most important prey item in Santa Cruz province, comprising 50% or more of puma diet in the two sheep ranches and Monumento Natural Bosques Petrificados, and 59% of consumed biomass in Torres del Paine National Park. In north-western Patagonia the guanaco occurs at a density of only 0.67 individuals per km2 and Novaro et al. (Reference Novaro, Funes and Walker2000) did not detect the species in the diet of the puma.
Among the other native mammals in the puma’s diet in Santa Cruz province edentates, present in the diet in Monumento Natural Bosques Petrificados, is a frequent puma prey item in other areas of Patagonia (Branch et al., Reference Branch, Pessino and Villareal1996; Novaro et al., Reference Novaro, Funes and Walker2000; Pessino et al., Reference Pessino, Sarasola, Wander and Besoky2001). The Patagonian mara is an uncommon item in the diet of pumas; it was found only in puma scats from the 25 de Marzo Ranch, which is at the southern limit of the distribution of the Patagonian Mara (Redford & Eisenberg, Reference Redford and Eisenberg1992). To our knowledge this is the first study to report its presence in the diet of the puma in an unprotected area. The contribution of rodent biomass to the puma diet in Santa Cruz was similar to that in north-western Patagonia (Novaro et al., Reference Novaro, Funes and Walker2000) but much less than in Central America, where rodents are the main prey item of the puma (Iriarte et al., Reference Iriarte, Franklin and Johnson1990a). This difference is probably due to the lack of larger prey in Central America, or because of trophic competition with the jaguar Panthera onca (Scognamillo et al., Reference Scognamillo, Maxit, Sunquist and Polisar2003).
Both the Magellanic penguin, in Monte León National Park, and the lesser rhea, in Monumento Natural Bosques Petrificados, were prominent in the puma’s diet. The Magellanic penguin is a newly recorded prey item for the puma. At Monte León National Park, Magellanic penguins nest under bushes, or dig burrows, throughout spring and summer (September–April; Schiavini et al., Reference Schiavini, Yorio, Gandini, Raya Rey and Boersma2005). The penguin can be a valuable prey for the puma not only because of its body mass (3.65 kg; Gandini et al., Reference Gandini, Frere and Holik1992) but also because of its abundance and predictability at the colonies. The second most consumed bird species, the lesser rhea, is distributed across the entire Santa Cruz province (Travaini et al., Reference Travaini, Bustamante, Rodriguez, Zapata, Procopio, Pedrana and Peck2007). This species was also consumed by pumas in Chile in Torres del Paine National Park but at a low frequency (Franklin et al., Reference Franklin, Johnson, Sarno and Iriarte1999).
Our results show that native prey, specially guanacos, are the most important prey for pumas in southern Patagonia, both in protected and unprotected areas, thus showing that native species are still performing an ecological function as prey for pumas. Our results do not support the suggestion of Novaro et al. (Reference Novaro, Funes and Walker2000) that native prey no longer have an ecological role as prey of the puma in Argentine Patagonia, at least in Santa Cruz province. Many sheep ranches in north and central Santa Cruz province were abandoned during 1991–2003 because of a combination of economic and climatic factors (Travaini et al., Reference Travaini, Zapata, Martínez-Peck and Delibes2000) and this could have lead to a resurgence of native prey. We suggest, therefore, that native prey could be restored to those areas of Argentine Patagonia, such as the north-west, where they are currently ecologically extinct. Facilitating native species recovery and/or restoration and applying more rigorous controls to prevent the introduction of potential alien prey species of the puma both, within and outside protected areas, needs to be evaluated as a regional strategy.
Acknowledgements
Funds were provided by the Universidad Nacional de la Patagonia Austral, Universidad Nacional de La Pampa, project PI29/B099 and the BBVA Foundation through a grant under the Conservation Biology Program. We acknowledge the support and permits provided by the National Parks Administration of Argentina and the ranch owners. During the writing of this article JIZM was supported by a Conservation, Research and Education Opportunities fellowship programme and Consejo Nacional de Investigaciones Científicas y Técnicas predoctoral fellowship. C. Zoratti, L. Montenegro, G. Soria, F. Escobar, G. Aguilera, P. Collavino, E. Daher, M. Yaya and M. Bronfman collaborated in the fieldwork. We also thank J.B. Mesa, J. Feaga and an anonymous reviewer for help and comments.
Biographical sketches
Juan Ignacio Zanón’s research focuses on the ecology and conservation of carnivores in Argentine Patagonia. Alejandro Travaini, Sonia Zapata and Diego Procopio are scientific researchers at the Research Center of Puerto Deseado–National University of Patagonia Austral. They are working on behavioural ecology and conservation of wildlife in the southernmost continental province of Argentine Patagonia. Miguel Ángel Santillán carries out research on raptors and passerines in La Pampa province of Argentina.



