


Introduction
In present-day Europe two members of the genus Canis, the wolf (C. lupus) and the jackal (C. aureus), exist. A similar situation prevailed during much of the Quaternary (e.g. Brugal & Boudadi-Maligne, Reference Brugal and Boudadi-Maligne2011), but there has been an ongoing debate on the moment of arrival of the first Canis. In September 2012 the annual Kor & Bot expedition dredging for fossils in the Oosterschelde estuary resulted, among numerous other finds, in the discovery of a fragmented canid mandible. The Oosterschelde (also known in literature as Eastern Scheldt) is a dammed-off estuary located in the province of Zeeland (the Netherlands) where fossiliferous submarine sediments have been dredged since the 1950s (De Vos et al., Reference De Vos, Mol, Reumer, VanKolfschoten and Gibbard1998; Scager, Reference Scager2015). The faunal content of the mammalian thanatocoenosis was published by (among others) De Vos et al. (Reference De Vos, Mol, Reumer, VanKolfschoten and Gibbard1998) and is now considered to date to the Early Pleistocene, MN17b sensu Palombo & Valli, Reference Palombo and Valli2003–Reference Palombo and Valli2004; MNR2 sensu Mayhew et al., Reference Mayhew, Dieleman, Slupik, van den Hoek Ostende and Reumer2014, c. 2.35 – 2.10 Ma (see Scager, Reference Scager2015). These fossils were subsequently resedimented in a Late Pleistocene channel lag deposited by the proto-Schelde (Slupik et al., Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, van Strydonck, Kiden, Burger and Reumer2013) and are a rare example of a reworked and ex situ faunule that is nonetheless considered as a faunistic entity.
Fossils are fished from the bottom of two deep gullies in the estuary, the Flauwerspolder pits 1 and 2 (formerly called Olifantenputje and Gastenputje; Scager, Reference Scager2015), both at close distance to the reinforced coast of the island of Schouwen-Duiveland. The fishing depth is c. 35–40 m (for the techniques used see Mol et al. (Reference Mol, van den Bergh and de Vos1999) and Reumer et al. (Reference Reumer, Mayhew and van Veen2005)). The sediment at that depth lies at the top of the Maassluis Formation and comprises medium- to fine-grained arenite with a small amount of lithic grains or with small particles of clay, manganese and shell fragments (Slupik et al., Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, van Strydonck, Kiden, Burger and Reumer2013).
The mandible fragment was easily recognised as belonging to a canid and has now been studied as it immediately raised questions concerning the relationship between the date of the faunule and the supposedly synchronous ‘Wolf Event’ (e.g. Sardella & Palombo, Reference Sardella and Palombo2007; hereafter WE). Carnivores from the Villafranchian Oosterschelde faunule had previously been restricted to Hyaenidae (Pliocrocuta perrieri) and Machairodontidae (Homotherium sp.; Scager, Reference Scager2015). The find of a canid mandible thus evoked considerable interest. The purpose of this paper is to identify the fragment, provide its description and check its probable relation to the Early Pleistocene WE. The WE was meant to mark the first appearance of the genus Canis in Europe (see Lacombat et al., Reference Lacombat, Abbazzi, Ferretti, Martínez-Navarro, Moullé, Palombo, Rook, Turner and Valli2008), in the form of the early wolf C. etruscus, c. 2.0 Ma ago (Cherin et al., Reference Cherin, Bertè, Rook and Sardella2013). However, quite a few taxa have been described within the genus, the exact status of which is open to debate. In addition, members of the genus were found in sediments of much older date.
Material and methods
The material was collected using a trawl net designed for commercially trawling edible mussels (Mytilus edulis). For this purpose, the vessel ZZ10 engages in a yearly traditional trawling expedition on the Oosterschelde estuary, trawling in the c. 35–40 m deep gullies south of the island of Schouwen-Duiveland. The collected bone and dental material is then desalinized (if necessary), conserved and stored. The mandibular fragment studied here consists of part of the left horizontal ramus with two premolars (p1 and p2), part of the canine (c) and part of the third premolar (p3) (see Fig. 1). The mental foramen is visible. It is stored at the National Museum of Natural History (NCB Naturalis), Leiden, under registration number RGM631800. Measurements were taken with a Vernier caliper, the photographs with a Canon Digital IXUS 65 camera.

Fig. 1. RGM631800; A. Buccal view; B. Lingual view; C. Dentition in occlusal view. Bar = 10 mm.
In order to identify the mandible fragment, comparative material was used from several collections in addition to published data. We studied material at NCB Naturalis, Natuurhistorisch Museum Rotterdam and the Laboratory of Paleontology (Department of Geology, Aristotle University of Thessaloniki) (see Table 1), and used published data from Del Campana (Reference Del Campana1913), Kerkhoff (Reference Kerkhoff1992), Rook (Reference Rook1993), Baryshnikov & Tsoukala (Reference Baryshnikov and Tsoukala2010) and Baryshnikov (Reference Baryshnikov2012).
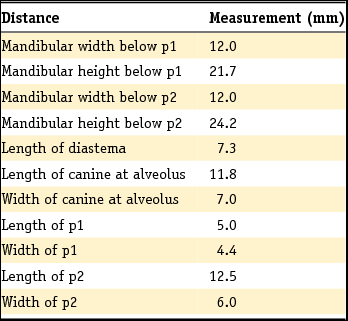
Table 1. Measurements for RGM631800.
Systematic palaeontology
ORDER Carnivora Bowdich, 1821
FAMILY Canidae Fischer von Waldheim, 1817
TRIBE Canini Fischer von Waldheim, 1817
GENUS Canis Linnaeus, 1758
Canis cf. C. etruscus Forsyth-Major, 1877
Material: RGM631800 (Fig. 1).
Locality: Oosterschelde (Zeeland, the Netherlands).
Age: Early Pleistocene (c. 2.4–2.08 Ma).
Description: This partial mandible has two complete premolars (p1 and p2), the anterior part of p3 and the canine (c), which is broken on the level of the alveolus. Moreover, the ramus is broken at the level of the canine on the anterior side and obliquely at the level of p3 on the posterior side. The rest of it is complete. The p1 is small and peg-like, and possesses one root. The p2 has a triangular shape in lateral view, with the major cusp (protoconid) situated at about two-fifths from the anterior. The posterior part of p2 has a ridge that ends in a hook shape without bearing a clear cuspid; the tooth has two roots. In occlusal view, it has the greatest width at the posterior half. There is one mental foramen beneath the anterior root of p2. The measurements of this specimen are given in Table 1.
Comparison: Unfortunately, our mandible only possesses premolars, which usually show intraspecific variability in their morphology and are therefore somewhat less diagnostic. The studied mandible fragment points to a robust large-sized canid. The basic difference between C. lupus, C. familiaris and the subgenus Canis (Xenocyon) is that these three taxa have one extra cuspid on p2, situated somewhat above the lower part of its posterior end. Similarities in the dental morphology exist with C. arnensis, C. etruscus, C. apolloniensis and C. mosbachensis. However, there is a significant size difference with the small-sized C. arnensis and C. apolloniensis, which is indicated not only by the dentition, but also by measurements of the ramus. Moreover, p1 of C. arnensis and C. apolloniensis are more elongated (length >> width) when compared to the p1 of C. etruscus and C. mosbachensis, which tend to be more semicircular (length > width). In addition, the small-sized specimens are considerably more slender than RGM631800.
Cherin et al. (Reference Cherin, Bertè, Rook and Sardella2013) published a revised diagnosis for the species C. etruscus. Most characteristics concern the skull and the upper dentition. Of the lower dental features mentioned in the revised diagnosis (‘Lower dentition characterized by larger dimensions in respect to C. arnensis and by a wolf-like m1/m2 ratio. The lower carnassial [m1] is distinguished by main trigonid cusps [protoconid and paraconid] relatively small [in comparison with C. lupus] and by talonid cusps [hypoconid and entoconid] linked by a sinous crest’) only the size can be used to compare our fragment. Nevertheless, all available characters point either to C. etruscus or to the only slightly smaller but (according to Brugal & Boudadi-Maligne, Reference Brugal and Boudadi-Maligne2011) conspecific C. mosbachensis. The dental dimensions show our mandible to be closer to C. etruscus. Moreover, the height and width of the ramus measurements also indicate a close relation to C. etruscus. Because of the presence of only the less diagnostic premolars, we tentatively attribute mandible fragment RGM631800 to Canis cf. C. etruscus.
Discussion
Brugal & Boudadi-Maligne (Reference Brugal and Boudadi-Maligne2011) recently revised and simplified the taxonomy of the Pleistocene members of the genus Canis by the recognition of only two species: (1) the small-sized C. arnensis (of which C. accitanus and C. apolloniensis are considered synonyms) and (2) the large-sized C. etruscus (which includes C. mosbachensis). This revision is mainly based on the fact that the two species groups are morphologically similar, differing only in size and size variability, which is not a factor of phyletic differentiation. By accepting this revision, the examination of our material indicates an identification as C. etruscus. The length of p1 falls in the C. arnensis group, although due to the length–width difference mentioned above, it has a closer morphologic relation to C. etruscus. In the case of p2, its size seems to fall in the C. etruscus group, with the C. arnensis group showing a clear overlap, although the latter is generally somewhat smaller. Again, the intraspecific variability of the premolars hampers a more precise conclusion.
The mandible represents the oldest find of the genus Canis in the Netherlands and surrounding regions of northwestern Europe. The fauna of the Oosterschelde of which it forms part is considered to have an age of about 2.35–2.10 Ma (De Vos et al., Reference De Vos, Mol, Reumer, VanKolfschoten and Gibbard1998; Scager, Reference Scager2015), although it is recovered from sediments dating to c. 30 ka due to resedimentation (Slupik et al., Reference Slupik, Wesselingh, Mayhew, Janse, Dieleman, van Strydonck, Kiden, Burger and Reumer2013). The age of the jaw fragment is supposedly 2.35–2.10 Ma and so it predates the published ages of the ‘Wolf Event’, which is supposed to have occurred during the early Pleistocene late Villafranchian, around 1.9–1.8 Ma ago (Azzaroli, Reference Azzaroli1983; Azzaroli et al., Reference Azzaroli, de Giuli, Ficcarelli and Torre1988; Sardella & Palombo, Reference Sardella and Palombo2007) or even around 1.4 Ma (e.g. Bonifay, Reference Bonifay and Lindsay1990). Along with a small-sized jackal-like Canis in the Mediterranean region (C. arnensis) appeared a larger wolf-like form, C. etruscus in the rest of the European continent. Younger forms of this taxon (often named C. mosbachensis) later gave rise to the wolf C. lupus (Garrido & Arribas, Reference Garrido and Arribas2008).
Other evidence seems to suggest, however, that the genus Canis first appeared in Europe as early as the Late Miocene (c. 7 Ma) in the Teruel Basin, eastern Spain (C. cipio, see Pons-Moya & Crusafont Pairo, Reference Pons Moya and Crusafont Pairo1978; Sotnikova & Rook, Reference Sotnikova and Rook2010). C. cipio is still a questionable taxon due to its scarce record, as only one fragmented maxilla and an isolated tooth were found; it is suggested not to belong to the genus Canis sensu stricto (Sotnikova & Rook, Reference Sotnikova and Rook2010). Another and more reliable early appearance of Canis (Canis sp.) is recorded from the site of Vialette, France, consisting of three fragmented mandibles, two isolated teeth and one tibia to which was attributed an age of c. 3.1 Ma (Lacombat et al., Reference Lacombat, Abbazzi, Ferretti, Martínez-Navarro, Moullé, Palombo, Rook, Turner and Valli2008). A canid (C. senezensis) was reported from from Senèze, France, dated to c. 2.0 Ma (Martin, Reference Martin1973; Roger et al., Reference Roger, Coulon, Thouveny, Féraud, Van Velzen, Fauquette, Cochemé, Prévot and Verosub2000) and a small canid (C. accitanus) from southern Spain, dated to c. 1.9 Ma (Garrido & Arribas, Reference Garrido and Arribas2008); both localities are slightly younger than the 2.35–2.18 Ma Oosterschelde find.
Looking back into the history of the early canids, it is clear that the story of the Canis lineage is complicated, especially in Europe. Because the attribution of C. cipio from the Late Miocene in Spain (7 Ma) to the genus Canis is doubted, the most probable hypothesis would be to situate the origin of the genus in North America at around 4 Ma, with its first appearance in Asia (East China) dated around 3.4 Ma (Flynn et al., Reference Flynn, Tedford and Qiu1991). Since the first accepted European Canini are dated around 3.1 Ma, it seems logical to conclude that Canis dispersed through Eurasia gradually and not instantaneously, and also much earlier than the Villafranchian. This confirms the doubts about the usefulness of the WE. The initial idea of this hypothesis was that the dispersal of Canis in Europe was accomplished in a short time interval, somewhere during the Villafranchian. We suggest here, following the suggestions of Rook & Martínez-Navarro (Reference Rook and Martínez-Navarro2010) and Sotnikova & Rook (Reference Sotnikova and Rook2010), that this dispersal lasted longer and must have been diachronous, hence not an ‘event’ but rather a process. It is not useful as a biostratigraphical marker.
It is also worth noting that the Oosterschelde find expands the geographical range of the genus Canis up to the edge of northwestern Europe, as the record until now spanned only up to central Europe (Brugal & Boudadi-Maligne, Reference Brugal and Boudadi-Maligne2011). This shows that the spatial dispersal of Canini was greater than currently believed.
Conclusion
The discovery of a Canis cf. C. etruscus mandible at the Oosterschelde locality (Zeeland, the Netherlands) shows that the genus Canis was dispersed up to northwestern Europe already sometime before 2 Ma, which expands its previously known geographical range. In addition, we agree that the so-called ‘Wolf Event’ cannot be used as a biochronological marker as the dispersal of the Canini through Eurasia occurred before this event and took place diachronously.
Acknowledgements
We thank the Schot family and the crew of the vessel ZZ10 for organising and managing the annual trawling trip in search of fossils. We thank Hans-Jorg Ahrens and Reinier van Zelst (NCB Naturalis, Leiden) for letting us study the mandible and giving us access to the collection of Naturalis. Professors Dimitris Kostopoulos and George Koufos (Laboratory of Paleontology, Aristotle University of Thessaloniki) are thanked for access to the Canis material stored in Thessaloniki. Wilma Wessels (Utrecht University) provided critical remarks, and reviewers Lorenzo Rook and John de Vos were of great help improving the initial manuscript.


