


Introduction
Lakes are a prominent feature of the ice-free regions of Antarctica and are also a major element of high-latitude northern landscapes. They are the downstream integrators of a variety of catchment processes, and many of their ecosystem properties are responsive to small variations in annual heat budget. These features, in combination with the strong climate-change signals and stratospheric ozone depletion at high latitudes, suggest thai polar lakes are highly appropriate sites for monitoring the effects of global change.
Recent advances in field instrumentation and remote-sensing technologies have paved the way for a variety of novel approaches to the monitoring of lake ecosystems. in this paper we examine some of the approaches which may be of special relevance to monitoring environmental change in the North and South Polar regions. We focus particularly on those properties of the aquatic environment which can be measured using optical techniques. The technology in this area is evolving rapidly and lends itself to high-resolution observations over space and time, even at high-latitude sites where the logistics are often difficult and expensive.
Long-term monitoring programs based on optical variables should ideally focus on those properties of lakes that not only respond to climatic forcing but also integrate a variety of lake-plus-catchment processes. Such variables should be measurable by in situ techniques, by automated instrumentation and/or by remote sensing. It would be advantageous if such a record could also be extended into the past, for example via paleolimnological approaches, to allow current observations to be placed in their historical context and to define the scales of natural variability against which recent changes can be assessed.
In this paper we examine several features of high-latitude lake ecosystems that are likely to respond to changes in climate forcing and which could be incorporated within a monitoring strategy based on optical properties: lake levels, ice-cover dynamics, phytoplankton biomass and chromophoric dissolved organic matter (CDOM). We place special emphasis on GDOM because it plays a central role in several ecosystem processes and is dependent on catchment hydrology, vegetation and soils, and thus climate. It exerts an over-riding control on underwater ultraviolet radiation (UVE) as well as photosynthctically active radiation (PAR) and provides a strong optical signature for long-term monitoring. Our results presented here show that high-latitude lakes have CDOM concentrations within a range where small variations result in biologically significant shifts in underwater UV-Α (UVR in the range 320-400 nm) as well as UV-B (280-320 nm). Such effects contrast with those induced by stratospheric ozone depiction which are limited to changes in UV-B, a waveband that penetrates to much shallower depths than UV-A in natural waters.
Lake Levels
One of the likely consequences of climate change is an alteration in the hydrological balance of lakes and their surrounding catchments. For example, a long-term warming trend in the boreal forest zone of Canada has been accompanied by reduced precipitation, decreased runoff and increased hydraulic residence time of the lakes of that region (Reference Schindler, Curtis, Parker and StaintonSchindler and others, 1996). For glacier-fed endo-rheic lakes, including those at many high-latitude sites, changes in the radiation balance at the glacier face can lead to large shifts in the quantity of inflowing meltwater and thus variations in lake area, depth and volume. Reference Chinn, Green and Fried mannChinn (1993) showed that lakes in the McMurdo Dry Valleys have consistently risen in water level throughout the 20th century, while measurements by Reference Gibson and BurtonGibson and Burton (1996) at lakes in the Vestfold Hills (East Antarctica) indicated a more complex pattern, with rising water between 1978 and the mid-1980s, followed by dropping water levels. These latter authors noted that variations in water level in saline meromictic systems are accompanied by changes in the vertical structure of the water column which in turn can be used to infer past climates. Given the large differences in reflectance between land, water and ice, changes in lake level could be conveniently monitored by aerial photography or by satellite, in particular via the associated changes in lake area. Alternatively, depth could be determined directly via airborne LIDAR (light detection and ranging) observations which can be used to estimate, under ideal conditions, shallows up to about 40 m deep (Gauldie and others, 1996).
Lake-Ice Dynamics
The dates of ice freeze-up and break-up, ice thickness and duration are all likely to be sensitive indicators of variations in climate. These variables will also have a wide-ranging influence on many physical and chemical properties of the aquatic ecosystem that directly or indirectly affect the biota, for example through changes in the length of the growing season, wind-induced mixing, gas transfer and underwater light availability. Reference WalshWalsh (1995) summarized the evidence for changing ice dynamics at a number of lake sites, and drew attention to the untapped potential of this approach to large-scale climate monitoring. Lake ice-cover thinning has been observed in continental Antarctica, suggestive of a warming trend, while measurements in the eastern Canadian Arctic show an increased frequency of multi-year lake ice (Reference Doran, McKay, Adams, English, Wharton and MeyerDoran and others, 1996) consistent with a cooling trend in this region.
Ice measurements can be made photographically (e.g. by automated cameras) or by sensors on aircraft or satellites (e.g. the advanced very high resolution radiometer). These optical approaches are likely to be superseded by satellite observations using synthetic aperture radar (SAR) based on active microwave sensing. SAR systems are continuing to improve in spatial resolution (30 100 m for RADAR-SAT) and they can be used to establish ice-cover extent even at night or under thick cloud cover ( Reference MassomMassom, 1995). SAR data have been successfully applied to measure the duration of ice cover and other surface ice properties of lakes in the Yukon and northern Alaska (Morris and others, 1995) and in the subarctic region near Hudson Bay (unpublished data from C. R. Duguay and P. M. Lafleur).
Phytoplankton
The potential for major shifts in the distribution and productivity of plant communities has been of particular interest to terrestrial ccologists in their development of climate-change scenarios for high-latitude environments. Various indices of greenness have been applied to some high-latitude environments, most recently by Reference Myneni, Keeling, Tucker, Asrar and NemaniMyneni and others (1997) who presented evidence of a 10 year consistent rise in the Normalized Difference Vegetation Index for latitudes higher than 45° N. Climate change is also likely to result in changing nutrient export from catchments, changes in the hydraulic residence time of lakes, and variations in lake-water mixing regimes, all of which may influence the species composition and biomass of lake phytoplankton.
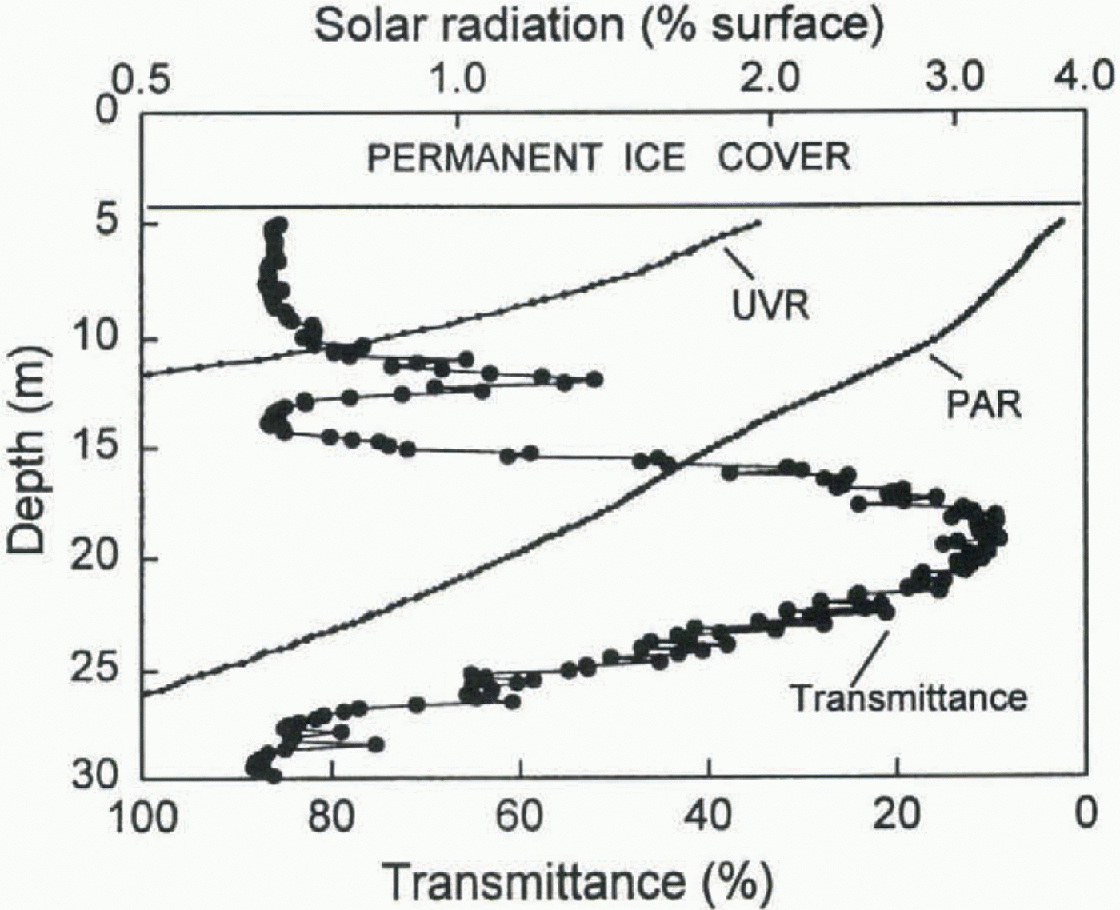
Fig. 1. Bio-Optical profiles for the east lobe of Lake Ronney, McMurdo Dry Valleys, Antarctica, 14 December 1995 (1215h): downwelling radiation at 320nm (UVR), down-welling photosynthetically active radiation (PAR) and transmittance at 660 ran (note the scale reversal to show the upper and lower pigment maxima). Measurements were made with a Biospherical PUV-500 profiling radiometer fitted with a Seatech 10 cm transmissometer.
A broad range of optical technologies is now available for rapid in situ measurements of phytoplankton distribution and abundance. High-resolution measurements of particle distribution can be made by fluorometry or trans-missometry which allow subsurface pigment maxima to be accurately located and quantified (Fig. 1). Techniques such as solar-induced fluorescence can provide information about the photosynthetic activity of phytoplankton as a function of depth in the water column (e.g. Reference Garcia-Mendoza and MaskeGarcia-Mendoza and Maske, 1996), and have been applied to lakes in both the North and South Polar zones (Vincent and others, in press).
A variety of remote-sensing options are available for phytoplankton measurements, and the advent of new satellite observation systems with high spatial resolution, such as MERIS and MODIS with resolution of 250-300 m (Massom, 1995), is of special interest to limnologists. Recent advances in laser fluorosensors indicate that LIDAR has considerable potential for airborne measurements of Chi a in the surface waters of lakes and coastal environments (Reference Nieke, Router, Heuermann, Wang, Babin and ThcrriaultNieke and others, 1997b). There is a great variety of paleo-limnological techniques based on the measurement of fossil algal remains in lake sediments (Reference Charles, Smol, Engstrom, Loeb and SpatieCharles and others, 1994 and references therein), and these approaches will continue to provide new insights into the long-term variability of high-latitude lakes (e.g. Pienitz and Smol, 1993).
Cdom
Climate-related changes in the hydrology and vegetation ecology of catchments are likely to have a strong effect on the export of dissolved organic compounds into receiving streams and lake waters. There is increasing evidence that such compounds exert a major influence on many properties of lake ecosystems, including radiative transfer (Laurion and others, 1997), mixing regimes (Reference Fee, Hccky, Kasian and CruikshankFee and others, 1996) and microbial carbon dynamics (Reference WetzelWetzel, 1995). in the Experimental Lakes Area a decrease in runoff and increased hydraulic residence time were accompanied by a 15-20% decrease in dissolved organic carbon (DOC) over 20 years (Schindler and others, 1996). The coloured fraction of the dissolved orgànics, referred to as CDOM, largely controls the spectral penetration of UVR and, to a lesser extent, pholosynthelically available radiation (PAR). in this section we evaluate the magnitude of these effects in high-latitude lakes, and the potential for monitoring coupled lake-catchment systems via the optical changes induced by variations in CDOM.
CDOM effects on underwater UVR
UVR penetrates deeply in high-latitude lakes as a result of the low concentration and low UVR-absorbing characteristics of CDOM in these waters Laurion and others (1997). Measurements of underwater spectral irradiance along latitudinal transects in northern North America show a major increase in the UV transparency of lakes across the transitional forest-tundra region of the sub-Arctic, and a further abrupt increase across the transition from tundía to polar desert (Vincent and Picnitz, 1996; Pienitz and others, 1997a, b; Vincent, in press). These changes are associated with the trend of decreasing concentration of CDOM with increasing latitude. Antarctic lakes are similarly characterized by low CDOM and the deep penetration of UVR (Fig. 1). High-latitude lakes are therefore little protected from the effects of stratospheric ozone depletion and rising UV-B, which are proceeding most rapidly in the polar regions.
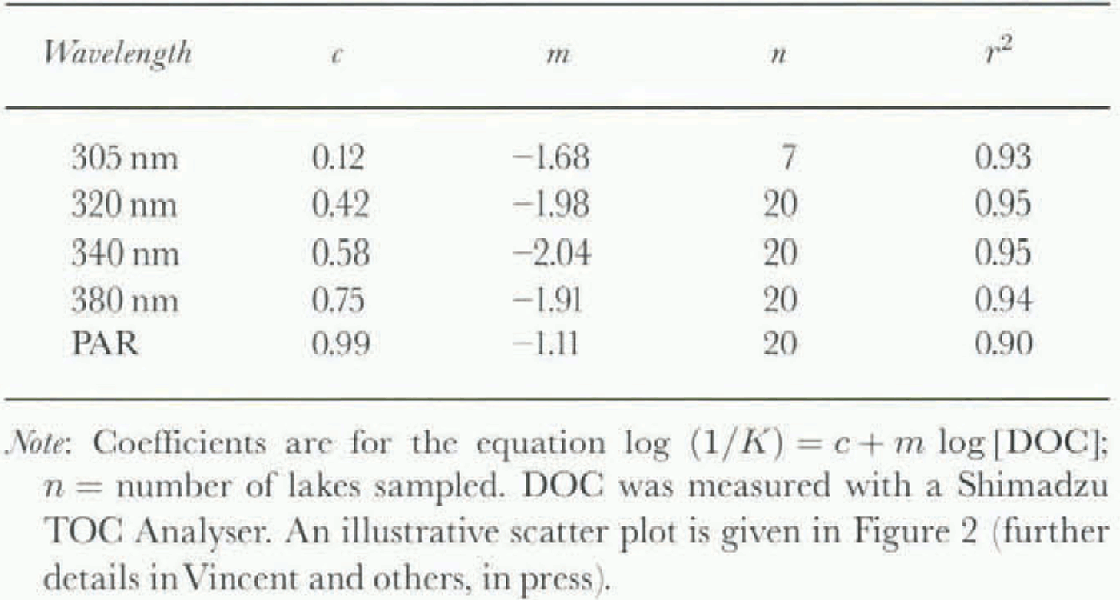
Our previous measurements of UVR in Arctic, sub-Arctic and Antarctic lakes have been made with a profiling radiometer (model PUV-500 from Biosphcrical Instrument Inc.) that measures downwclling LIVR at 305, 320, 340 and 380 nm. From these measurements a set of empirical relationships was derived for attenuation length or “transparency” ![]() where Κ is the diffuse attenuation coefficient) as a function of DOC concentration (table 1). The striking goodncss-of-fit of this log-log model (Fig. 2) may in part reflect the lack of bio-optical diversity amongst the sampled lakes; all are oligotrophic, non-turbid systems in which seston (biotic and abiotic suspensoids) plays only a minor role in light absorption and scattering.
where Κ is the diffuse attenuation coefficient) as a function of DOC concentration (table 1). The striking goodncss-of-fit of this log-log model (Fig. 2) may in part reflect the lack of bio-optical diversity amongst the sampled lakes; all are oligotrophic, non-turbid systems in which seston (biotic and abiotic suspensoids) plays only a minor role in light absorption and scattering.
Table. 1. Empirically derived relationships between attenuation length (1/K) and DOC concentration Jar Arctic, sub-Arctic and Antarctic lakes
Wc can extend these empirical transparency models by considering the penetration of UVR weighted for its biological effectiveness and for the changes in incident spectral radiation that result from ozone depletion. A parameter Τ*(λ), the “weighted transparency” at each wavelength, may be defined as:
where Κ is the diffuse attenuation coefficient at that wavelength (λ), ![]() is the biological weighting function which expresses the damaging effect of radiation at that wavelength on a relative scale (
is the biological weighting function which expresses the damaging effect of radiation at that wavelength on a relative scale (![]() is the surface irradiance at that wavelength, also on a relative scale (
is the surface irradiance at that wavelength, also on a relative scale (![]() , and F is the factor of enhancement in surface radiation flux at the wavelength for a given stratospheric ozone depletion. This weighted transparency
, and F is the factor of enhancement in surface radiation flux at the wavelength for a given stratospheric ozone depletion. This weighted transparency
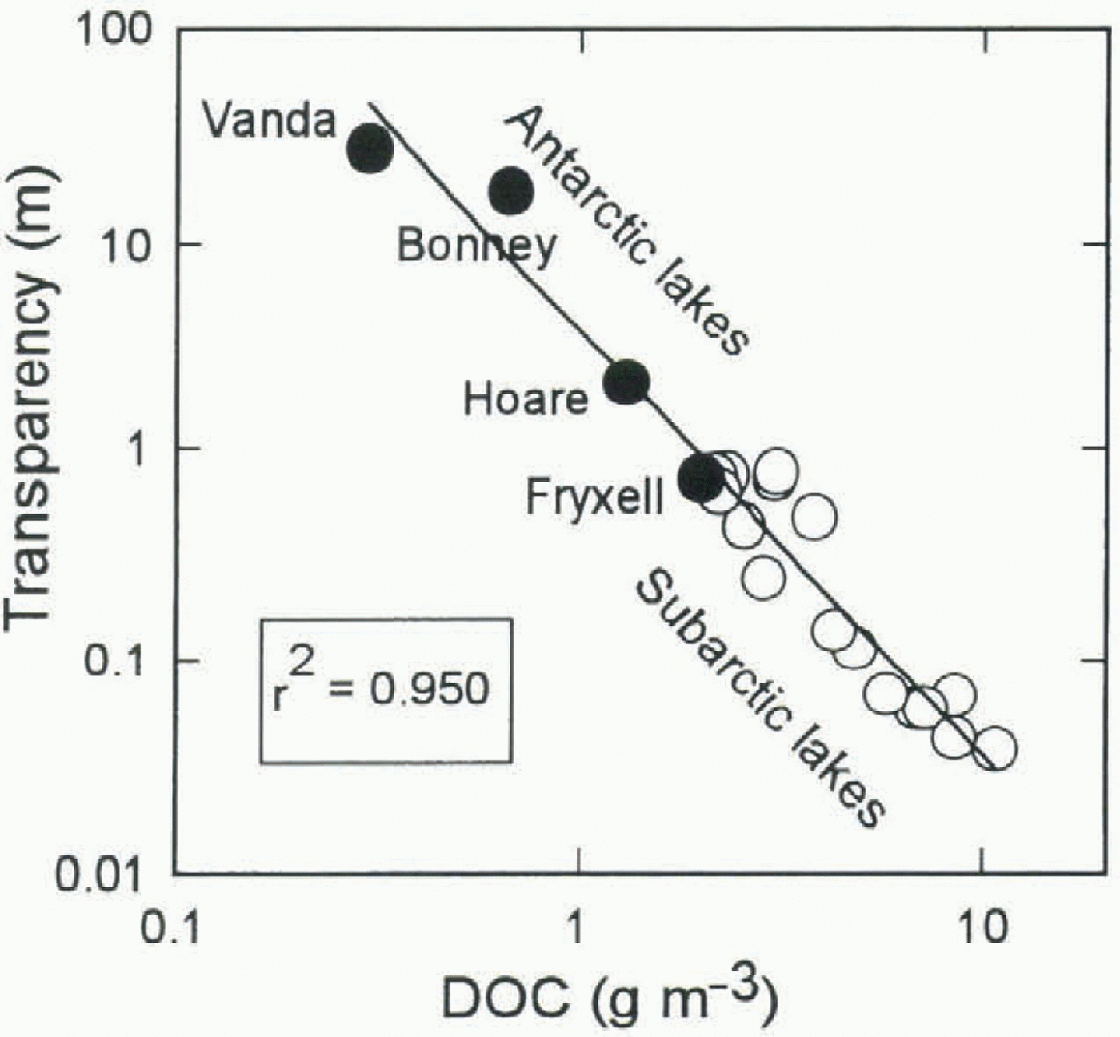
Fig. 2. Relationship between UVR transparency (1/K(λ). here illustrated by λ = 340nm) and DOC concentration for Antarctic lakes (solid circles) and sub-Arctic lakes (open circles).
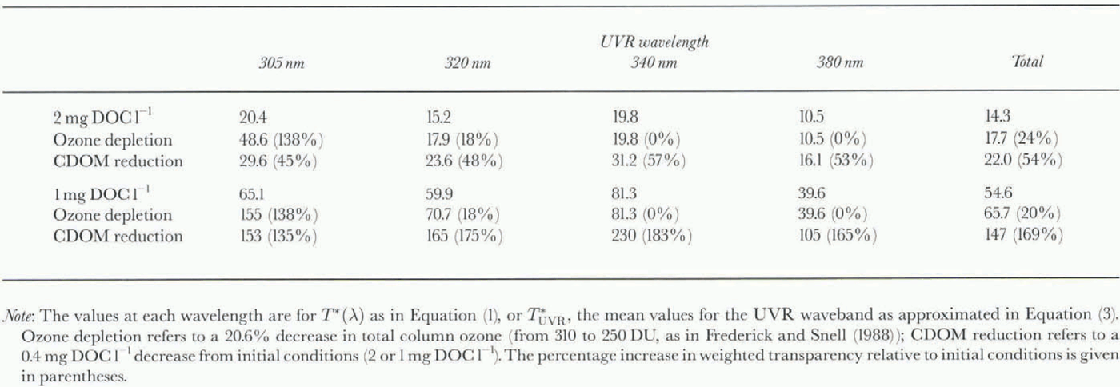
Table. 2. Effect of ozone depletion vs CDOM reduction on the weighted transparency (10−3 m) of a water column to UVR
index ran be averaged across a series of wavelengths to produce a mean value for a particular waveband:
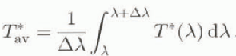
T*av ihr the UVR waveband can be approximated in terms of
![]() values for each of the lour measured wavelengths:
values for each of the lour measured wavelengths:
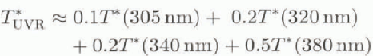
where the coefficients in this equation are for rectangular integration across the UVR range.
Table 2 presents the results of a series of calculations based on Equations (1) and (3) and the bio-optical relationships for high-latitude waters given in Table 1. For these calculations we used e values based on Reference Cullen, Neale and LesserCullen and others (1992). These were derived for UVR photo-inhibition of photosynthesis of marine diatoms, but they have also provided reasonable biological weightings lor UVR inhibition of growth by Antarctic cyanobacteria (Reference Quesada and VincentQuesada and Vincent, 1997). Eorel values were obtained for an incident solar radiation curve (Reference Milot-Roy, Vincent, Williamson and ZagareseMilot-Roy and Vincent, 1994, Fig. 7), and F values were derived from Reference Frederick and SnellFrederick and Snell (1988, fig. 2B) for a total column ozone thickness of 310 and 250 Dobson Units (DU).
Relative to the initial conditions of 2 mg DOCl1 and 310 DU, a 20.6% reduction in ozone results in a >100% increase in Τ* (λ) at short UVR wavelengths, but no change in long-wavelength UVR (table 2). By contrast, a 20% decrease in DOC from 2 to 1.6 mg Cl−1 results in a large (>40%) increase in Τ*(λ) at all UVR wavelengths, and the relative increase in total waveband ![]() is twice that induced by the ozone depletion. These effects are more striking at lower DOC concentrations. For example, the same absolute change in DOC, but from 1 to 0.6 mgl−1, results in a >130% increase
is twice that induced by the ozone depletion. These effects are more striking at lower DOC concentrations. For example, the same absolute change in DOC, but from 1 to 0.6 mgl−1, results in a >130% increase ![]() at all UVR wavelengths; T*UVR
increases by 169%, an effect some 8 times greater than that caused by 60 DU of ozone depletion (table 2).
at all UVR wavelengths; T*UVR
increases by 169%, an effect some 8 times greater than that caused by 60 DU of ozone depletion (table 2).
This analysis of weighted UVR penetration into natural waters illustrates the pivotal role played by CDOM and is consistent with earlier suggestions by Schindler and others (1996) and Reference Williamson, Stemberger, Morris, Frost and PaulsenWilliamson and others (1996) of the importance of such effects relative to changes in incident UVR. Our results show that in the low-CDOM regime which characterizes high-latitude lakes, changes in the concentration of DOC can have a greater effect on biologically weighted underwater UVR than the equivalent percentage change in stratospheric ozone. Much of this effect is due to UV-A, because the reduced photodamaging effect per unit energy (i.e. lowe ![]() ) in this waveband is offset by its deeper penetration and higher incident flux at the lake surface relative to UV-B.
) in this waveband is offset by its deeper penetration and higher incident flux at the lake surface relative to UV-B.
CDOM effects on underwater PAR and UVR/PAR
Although CDOM absorbant e of solar radiation is especially strong in the UVR region of the spectrum, there is also some absorbance in the PAR range. Our measurements in high-latitude northern lakes indicate that CDOM can account for a large percentage of the total photon budget for PAR and that climate-related shifts in DOC export to lakes could also influence the availability of underwater light for photosynthesis (unpublished data from S. Marka-ger and W. F. Vincent). Reference Arrigo and BrownArrigo and Brown (1996) modelled the combined effects of CDOM on UVR screening and PAR removal in the sea, and concluded that the latter can eliminate or exceed the positive elfccts of the former. For a decrease in CDOM from 2.0 to 1.6 mg DOCl−1, as above, wc calculate that the transparency of the water column to PAR would increase by only 11 %, but a shift from 1 to 0.6 mgl−1 would increase transparency (e.g. the depth of the 1 % PAR level) by 76%.
UVR impairment of biological systems is likely to represent the net balance between photochemical damage and biosynthetie repair. The latter is determined in part by the UVR flux, while biosynthesis in photoirophic organisms may depend on PAR availability. The spectral ratio of UVR to PAR may therefore be a more meaningful index of damage potential than UVR alone. Studies on high-latitude lakes in the northern circumpolar region (Vincent and Pienitz, 1996; Laurion and others, 1997) as well as Antarctica (Vincent and others, in press) have shown these waters lie within a CDOM concentration range where this ratio rises sharply with decreasing DOC. For the two CDOM scenarios and the empirical relationships given in Tabic 1, the UVR (320 nm)/PAR ratio would rise by 22% for the shift from 2 to 1.6 mg DOCl−1, and by 56% for the shift from 1 to 0.6mgDOCl−1.
Optical monitoring of CDOM
The preceding subsections have emphasized how CDOM controls the underwater light field in high-latitude lakes. It follows that certain optical properties of these lakes can be used as sensitive proxies for CDOM, for example attenuation lengths derived from in situ UVR profiles (Fig. 1; Table 1). For remote-sensing applic ations, attention has especially focused on the spectral fluorescence properties of CDOM, in particular its emission maximum normalized to the Raman scattering peak for water (Creen and Blough, 1995; Nieke and others, 1997a). Figure 3 shows the emission spectra for two lakes at opposite ends of the CDOM continuum for high-latitude waters: Lake Bonney (Antarctica) in which CDOM emission is small relative to the Raman peak (DOC = 0.7 mgl−1), and a sub-Arctic lake characterized by much higher concentrations of CDOM (8 mg DOCl−1) in which the Raman peak is a small contribution to total fluor-escence. Emission signals of this type from surface waters can be detected and quantified by active remote-sensing techniques such as LIDAR (Vodacck and others, 1995; Nickc and others, 1997a). CDOM can be incorporated into the ice of northern lakes during freeze-up (R. Pienitz and W. F Vincent, unpublished observations) and the organic content of lake ice might also be monitored in this way, although such measurements would be obstructed by any overlying snow.
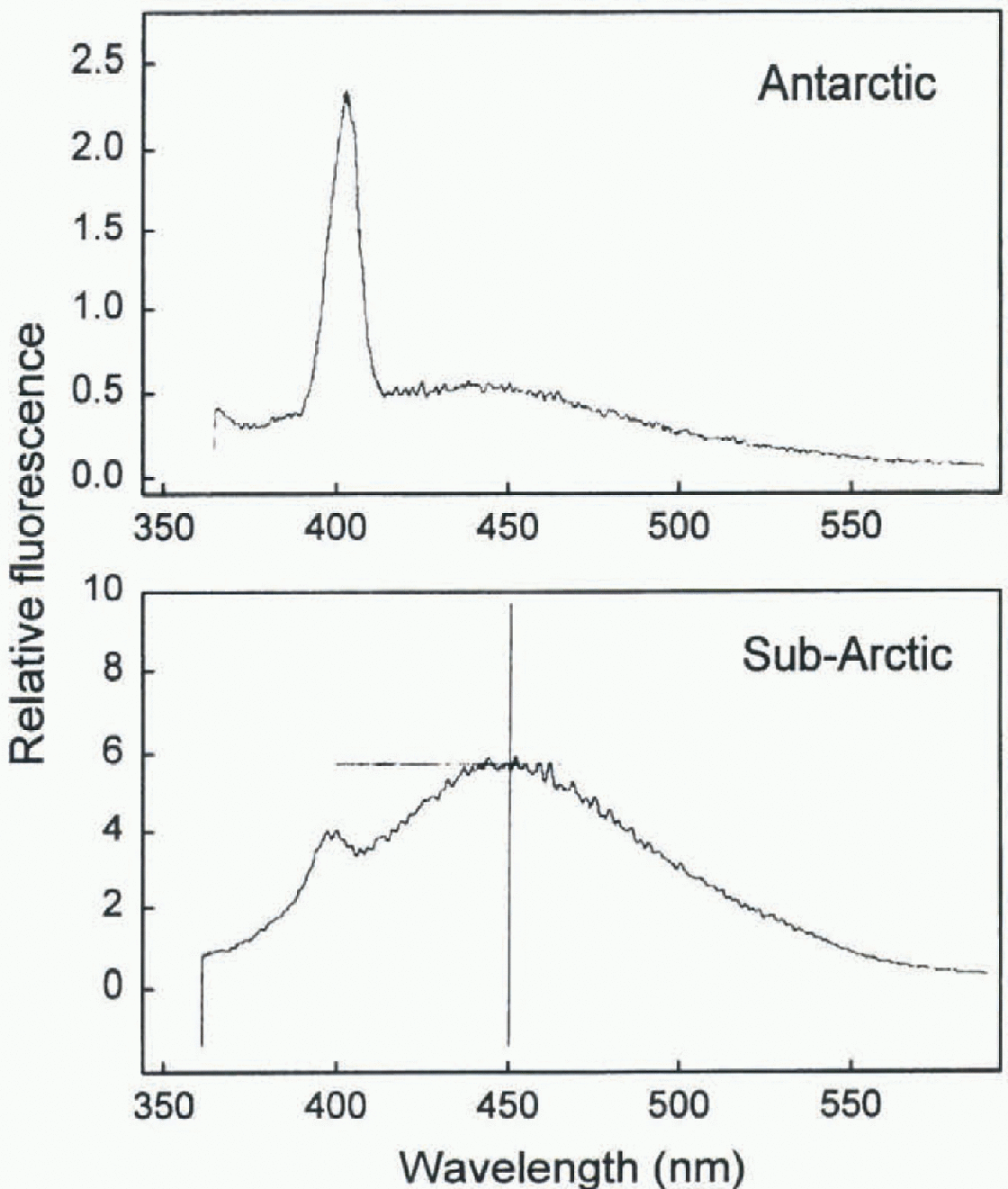
Fig. 3. Fluorescence emission spectra for 0.22μm filtered water from an Antarctic lake (Lake Bonney) and a sub-Arctic lake (near Lac à l'eau claire, northern Québec). Excitation was at 348nm, and measurements were obtained with a Shimadzu speclmfiuorometer model RF5000.
Recent advances in paleolimnology have led to the development of new analytical and statistical techniques for reconstructing past lake environments (Charles and others, 1994). These approaches are of Special interest for high-latitude lakes, where there is a paucity of historical data but a potentially interesting record of variations because of their high sensitivity to natural as well as human perturbation. Pienitz and Smol (1993) have shown that dissolved organic carbon levels in Canadian lakes can be successfully hindcast from the multivariate analysis of diatom microfossils contained with in their sediments. in combination with the bio-optical models now available for high-latitude lakes (table 1), this leads to the exciting prospect of reconstructing past underwater light regimes and the extension of records for optical variables such as T* into the past. This new theme in paleolimnology, “lake palco-optics” (Vincent and Pienitz, 1996), offers opportunities for integrating studies from the present-day optical monitoring of lakes, as proposed above, with knowledge of their historical variations in underwater light regime.
Acknowledgements
Ί his work was supported by grants from the Fonds pour la Formation de Chercheurs et l'Aide à la Recherche FCAR, Québec i and the Natural Sciences and Engineering Research Council (NSERC, Canada). Logistic support was provided by the United States Antarctic Research Program, Centre d'études nordiques and Polar Continental Shelf Project (this is publication No. PCSP/ÉPCP 01897). We thank K. Michael, J. C. Ellis-Evans and J. A. E. Gibson for their helpful review comments.





